DISKUSSION
Kommentare und Einwände von PD. Dr. S.L. [Initialen vgl. p. 570 Mitte. Der Autor gehört zu den führenden deutschsprachigen Evolutionsbiologen. Ich gebe seine Kommentare und Einwände hier wieder, weil ich sie für weitgehend typisch für das evolutionistische Denken halte und der Leser die unterschiedlichen Positionen in direkter Diskussion mit Gewinn verfolgen kann.], (Universität K.) (vollständige Wiedergabe) plus meine Anmerkungen:
S. L.: "Vielen Dank für das Buch "Artbegriff und Ursprung der Arten", das Sie mir Anfang Juli [1987] zugeschickt hatten. Ich bin erst jetzt dazu gekommen, es ausführlicher zu lesen.
Ausgesprochen gut gelungen ist das Thema, wieviele Arten von Lebewesen es gibt. Es ist völlig richtig: Die Antwort auf diese Frage hängt von der Annahme des jeweiligen Artbegriffs ab. Gekonnt zeigen Sie auf, dass verschiedene Autoren, selbst wenn sie gleichermaßen die Evolutionstheorie akzeptieren, zu unterschiedlichen Antworten auf die Frage kommen und sich zum Teil sogar selbst widersprechen. Ich bedaure, dass eine derartige Kritik nicht auch aus dem Lager der Evolutionsbiologen kommt.
Sehr gekonnt setzen Sie sich auch mit Beispielen auseinander, die immer wieder als Argument für den status nascendi von Arten aufgeführt werden. In der gängigen Evolutionsliteratur sind diese Beispiele in der Tat viel zu glatt dargestellt, und in der Tat hält man sich oft genug nicht streng genug an vereinbarte Definitionen.
Zu Recht betonen Sie auch, dass der Nachweis von Artaufspaltungen von großer Wichtigkeit ist für die Evolutionstheorie, nach der die ganze Artenvielfalt durch Artaufspaltungen und Bastardierungen entstanden sei."
W.-E. L.: Da möchte ich weiter differenzieren: Die Artaufspaltungen im Sinne des morphologischen Artbegriffs z.B. halte ich für die Kernfragen der Evolutionstheorie (Makro- und Megaevolution) wie dem Ursprung synorganisierter neuer Strukturen und Organe sowie der Baupläne im Tier- und Pflanzenreich für durchweg irrelevant. Das trifft genauso auf den neodarwinistischen und genetischen Artbegriff zu (zumindest kenne ich bis heute trotz intensiver Suche keine Ausnahme). Interessant wird die Sache erst beim genetisch-plasmastischen Artbegriff, der in etwa der morphologischen Abgrenzung zwischen den Triben (in der Botanik) und den Familien (öfter in der Zoologie) entspricht. Hier wird es auch mit morphologischen, genetischen und paläontologischen Übergangsformen schwierig (vgl. ARTBEGRIFF, 314 - 322). Für die oben erwähnten Kernfragen der Evolutionstheorie wären Artaufspaltungen durch Neubildung genetischen Materials notwendig (vgl. ARTBEGRIFF, p. 233, Zitat Vollmert und das Mutationskapitel p. 325 ff. - die in diesem Brief angegebenen Seitenzahlen beziehen sich von jetzt ab immer auf diese Arbeit ohne sie jedesmal zu nennen). Wo auf unserer Erde findet etwa die Bildung eines Oberschnabelgelenks bei Vögeln ohne eine solche Einrichtung durch richtungslose Genmutationen oder sonstwelche ungerichteten genetischen Veränderungen statt? (Siehe p. 264 f. und Fragen p. 268) Nirgends! Statt dessen versucht nun die neodarwinistische Evolutionstheorie mit einem enggezogenen Artbegriff - (eng, weil die entscheidende Frage nach neuer genetischer Information für den Ursprung synorganisierter Strukturen und Organe etc. darin gar nicht vorkommt) - Artaufspaltungen zu demonstrieren und mit solchen Artaufspaltungen den Modus des Ursprungs aller Lebensformen zu implizieren. Und auf solche Beispiele trifft darüberhinaus Ihre Bemerkung zu:
"In der gängigen Evolutionsliteratur sind diese Beispiele in der Tat viel zu glatt dargestellt, und in der Tat hält man sich oft nicht streng genug an vereinbarte Definitionen."
Weiter im Text:
S. L.: "Wenn Sie aber meinen, dass die ganzen von Ihnen aufgeführten Beispiele irrelevant für die Begründung der Evolutionstheorie seien, so kann ich Ihnen nicht zustimmen aus folgenden Gründen:
- Zunächst einmal sind die ganzen Beispiele verträglich mit der Evolutionstheorie."
W.-E. L.: Ich bringe ja nun serienweise Beispiele für Variabilität durch Strukturabbau (vgl. z.B. das Degenerationskapitel p. 403 f., aber auch 374 f. oder 122 f.). Die Evolutionstheorie wollte doch vor allem aber den Ursprung, den Aufbau der Information und Synorganisation im Bereich des Lebendigen erklären (vgl. p. 403, Zitat Chargaff); die Anwendung des Begriffs Evolution für den Abbau von Strukturen und Degenerationserscheinungen überhaupt ist ja ein ziemlich starker Bedeutungswandel der ursprünglichen Zielsetzung. Und historisch gesehen war die Anwendung dieser Begriffsveränderung (oder wenn Sie wollen: Begriffserweiterung) durch H.G. Wells (The Time Machine 1895) ein ziemlicher Schock für seine Zeitgenossen. Diese Begriffserweiterung hat bedauerlicherweise zur Begriffsverwirrung beim Evolutionsthema beigetragen:
An vielen Stellen spricht man von Evolution und impliziert dabei Strukturaufbau und bringt dann häufig Beispiele für Informations- und Strukturverlust als Evolutionsbeweise! Da ja alles Evolution ist, spielt das scheinbar keine Rolle mehr. "Dass vorhandene Strukturen durch Mutationen abgebaut, aufgelöst, total ausgelöscht werden können, ist eine altbekannte Tatsache. Aber noch einmal: Kann uns dieses Phänomen den Ursprung der Strukturen erklären? Hier werden doch zwei grundverschiedene Dinge miteinander verwechselt! Die Evolution soll uns den Ursprung der Ordnung und der integrierten Mannigfaltigkeit an Strukturen im Bereich des Lebendigen erklären und als Beweise werden uns Fälle vorgeführt, bei denen der Verlust der Ordnung... (und) an Strukturen empirisch konstatiert wird, wohl aber mit einem Selektionsvorteil verbunden ist. Der Selektionsvorteil erklärt jedoch nicht den Ursprung der Ordnung! Der Selektionsvorteil erklärt uns nur, warum bestimmte Formen unter gewissen Bedingungen noch existieren, aber nicht wie sie in ihrer ursprünglichen Form ins Dasein gekommen sind!" (Lönnig 1971, pp. 21/22 im Anschluss an die Beispiele der Sichelzellanämie und Inselbewohnern mit stummelförmigen Flügeln als Evolutionsbeweise nach Egelhaaf 1969).
Ich möchte Ihnen zustimmen: Die ganzen Beispiele sind verträglich mit der Evolutionstheorie - in ihrer erweiterten Fassung und vor allem mit dem Teil, der mit der ursprünglichen Zielsetzung wenig zu tun hat, dem Verlust von Information und Strukturen auf allen Ebenen: genetisch, physiologisch, morphologisch-anatomisch und ethologisch.
Wesentlich besser vertragen sie sich jedoch mit der Schöpfungslehre (ursprünglich vollkommene Schöpfung mit hohem Anpassungspotential und dann Degeneration).
S. L.: "Die Tatsache, dass es Rassen gibt, die sich noch kreuzen lassen und die sich dennoch gut unterscheiden, zeigt: Selbst beim Fehlen von genetischen Barrieren können sich verschiedenartige Populationen - eben die Rassen - herausbilden. Die Tatsache, dass in etlichen Fällen sogar schon eine genetische Barriere ausgebildet wurde, zeigt, dass eine Rassenbildung schließlich zur Existenz wirklich getrennter Populationen führen kann."
W.-E. L.: Das ist nicht zu bezweifeln. Der Abbau von Strukturen hat schon oft zur Rassenbildung (Musterbeispiel Hunde! - siehe p. 599, Zitat Trumler) und auch zu genetischen Barrieren zwischen Populationen geführt (vgl. 131 ff.). Besonders im Pflanzenreich liegt hierfür die Ursache auch häufig in der Verdoppelung bzw. Vervielfachung des Genoms (Polyploidie), aber auch Chromosomenmutationen haben das Ihrige im Tier- und Pflanzenreich beigesteuert (wobei die letzteren häufig wieder mit Verlust genetischen Materials einhergehen - vgl. p. 408). Ich bezweifle nur, dass solche Artbildung etwas mit der oben zitierten ursprünglichen Zielsetzung der Evolutionstheorie zu tun hat.
S. L.: " - Wenn eine Artaufspaltung noch nicht abgeschlossen ist, ist natürlich immer mit Bastardierungen zu rechnen."
W.-E. L.: Klar! Mich stört nur, dass in der evolutionistischen Literatur die Frage nach den Hybridisierungsmöglichkeiten oft gar nicht erst gestellt wird, man verschiedenen Populationen einfach verschiedene Artnamen gibt und dann das Ganze als Artbildungsbeweise vermittelt. Aber warum auch nicht! Es handelt sich dann um Arten im Sinne des morphologischen oder neodarwinistischen Artbegriffs und diese (und andere Artbildungsmodi) haben mit den Kernfragen der Evolutionstheorie nichts zu tun.
S. L.: "Nach der Evolutionstheorie werden u.a. Artaufspaltungen als notwendig für den Evolutionsprozess angesehen, und dass diese notwendige Bedingung offensichtlich zu allen Zeiten - so auch jetzt noch - gegolten hat und noch gilt, das also wird durch die Beispiele gut belegt, und deswegen sind sie auch verträglich mit der Evolutionstheorie. Wenn die Artbildung völlig abgeschlossen ist, sind Bastardierungen nicht mehr möglich."
W.-E. L.: Auch die häufig mit Strukturverlust einhergehende Artaufspaltung des genetischen Artbegriffs kommt an die Hauptprobleme der Evolutionstheorie nicht heran. Das habe ich in der Artbegriffsarbeit ausführlich diskutiert. Meiner Auffassung nach differenzieren Sie noch zu wenig beim Artbegriff und der Artaufspaltung. Wir kennen bisher keine Artaufspaltung durch systematischen Aufbau neuer genetischer Information (Serien von Struktur- und Regulatorgenen, entsprechend Aufbau neuer Substanzen und morphologisch-anatomischer Strukturen, Synorganisationen solcher zu neuen biologischen Systemen etc.). Solche Artaufspaltung sehe ich als notwendig für den Evolutionsprozess an. Und dass die bekannten Beispiele von Artaufspaltungen diese notwendige Bedingung heutzutage nicht durch die Faktoren der Synthetischen Evolutionstheorie erfüllen - und so auch zu keiner Zeit - , das macht sie so irrelevant für die Evolutionstheorie.
Im übrigen unterscheiden wir ja sehr klar zwischen notwendigen und hinreichenden Bedingungen. Die bekannten Beispiele sind für die zitierten Kernfragen der Evolutionstheorie weder notwendig noch hinreichend.
S. L.: "Daraus aber den Schluss abzuleiten, dass solche Arten nur durch einen intelligenten Schöpfungsakt entstanden sein soll, halte ich nicht für zwingend."
W.-E. L.: Ich auch nicht: Solche morphologischen, neodarwinistischen und durch Strukturabbau entstandenen genetischen Arten sind nicht durch einen intelligenten Schöpfungsakt (als direkten Eingriff Gottes in die Natur) entstanden. Sie sind auf Abbau, Umbau, Verdoppelung und Rekombination genetischen Materials zurückzuführen (vgl. auch p. 473). Nur Arten mit systematischem Gewinn an genetischer Information, mit Neukonstruktionen samt Synorganisationen auf den verschiedensten Ebenen sind durch einen intelligenten Schöpfungsakt entstanden (siehe pp. 210 ff., 283 ff., 323, 359, 470).
S. L.: "Allenfalls besteht eine Verträglichkeit der Beispiele mit dem Schöpfungsgedanken."
W.-E. L.: Sowohl 1. die primäre Artbildung (Aufbau) als auch 2. die sekundäre (Abbau) wird von der Schöpfungslehre als 1. ursprüngliche Schöpfung und Anpassung und 2. anschließende Degeneration postuliert (p. 473).
S. L.: "In einem solchen Fall würde sich aber fragen, warum die Intelligenz nur bei der Erschaffung von Arten die einer neuen Familie oder noch höheren taxonomischen Einheit angehören, wirksam sein soll und nicht bei der Erschaffung von Rassen und Induviduen."
W.-E. L.: Weil die Intelligenz für den Aufbau primärer Arten notwendig ist, nicht aber für den Zerfall in sekundäre Arten (der wohl z.T. programmgemäß abläuft - vgl. p. 144 ff.). Die Erschaffung von Individuen ist durch die Fortpflanzung mit Ontogenese gesichert - diese Programme sind mit den Arten erschaffen worden, nicht aber die Möglichkeit primäre neue Arten zu bilden. In einer Blaualge gibt es kein Programm für eine Kastanie und in der Spitzmaus kein Programm für die Entwicklung des Menschen, wohl aber in allen Organismen fein ausdifferenzierte Programme für deren Arterhaltung.
S. L.: "Berd-Olaf Küppers hat wiederholt darauf hingewiesen, dass die Zufallshypothese aus prinzipiellen Gründen unbeweisbar, und die teleologischen Hypothesen aus prinzipiellen Gründen unwiderlegbar seien (z.B. in Küppers 1987: Ordnung aus dem Chaos, p. 42). So gesehen kann ich Ihre Hypothese nicht widerlegen."
W.-E. L.: Zum zweiten Punkt bin ich anderer Auffassung: Der Ansatz der Erschaffung primärer Arten im zitierten Sinne ist empirisch widerlegt, wenn sich entweder herausstellt, dass in der Blaualge doch ein Programm für die Kastanie vorliegt und in der Spitzmaus ein Programm für die Bildung des Menschen etc., oder dass sich solche Programme (immer induktiv-mutationsgenetisch bewiesen und reproduzierbar) de novo und ungezielt bilden können. Beides ist nicht der Fall; vgl. dazu das Mutationskapitel. Alle Erfahrung lehrt uns vielmehr, dass synorganisierte Um- und Neukonstruktionen den Konstrukteur implizieren (Details pp. 282 - 290).
Im übrigen stellt Vollmer in der Besprechung von Küppers Buch DER URSPRUNG DER BIOLOGISCHEN INFORMATION (1986) in der Naturwissenschaftlichen Rundschau (40, 246/247; 1987) zu dessen molekulardarwinistischem Ansatz u.a. fest:
"Wenn Küppers kritisch feststellt, dass die Zufallshypothese nicht beweisbar, der teleologische Ansatz dagegen nicht widerlegbar sei (...), so muss doch ebenso kritisch festgehalten werden, dass der molekulardarwinistische Ansatz weder beweisbar noch widerlegbar ist, so dass auch seine eigentlichen Vorzüge anderswo liegen müssen. Hier wird anscheinend mit zweierlei Maß gemessen.
...Schließlich bleibt auch der molekulardarwinistische Ansatz vorerst jede Abschätzung über die Wahrscheinlichkeit der Lebensentstehung schuldig.
...Auch die zentrale Frage, wie es bei Organismen zur Neuentstehung von Genen kommen kann, bleibt völlig ausgeklammert."
Weiter zu Ihren Einwänden:
S. L.: "Aber Ihre Argumentation kann auch nicht die Evolutionstheorie widerlegen. Sie können allenfalls versuchen zu zeigen, dass bisherige Argumente nicht hinreichend sind zur Begründung der Evolutionstheorie, und selbst dann kommt es noch darauf an, innerhalb welches Axiomensystems man argumentiert."
W.-E. L.: Eine wissenschaftliche Theorie sollte in der Lage sein, klare Falsifikationskriterien zu nennen. Ich habe solche für meinen Ansatz oben genannt. In Hunderten von Gesprächen habe ich jedoch regelmäßig keine Antwort auf diese Frage für die Evolutionstheorie bekommen. Ich bin nun natürlich sehr gespannt, welche Falsifikationskriterien Sie mir für Ihre Theorie nennen werden!
Weiter zum Axiomensystem:
S. L.: "Es gehört zum Axiomensystem der gegenwärtigen Naturwissenschaft, nicht das Walten höherer Kräfte für die Erklärung natürlicher Phänomene zu benutzen. Sie tun es, und insofern besteht ein wichtiger axiomatischer Unterschied in den verschiedenen Argumenten."
W.-E. L.: Dieses Axiomensystem ist für viele zu einem Denkverbot zur Frage nach dem Konstrukteur der ungeheuer komplexen Konstruktionen in der Natur geworden.
Es ist ja gerade die Frage, ob Makro- und Megaevolution überhaupt "natürliche Phänomene" sind: solche Evolution ist jedenfalls nirgends zu beobachten und auch nach den Voraussetzungen der Synthetischen Evolutionstheorie nirgends mutationsgenetisch oder sonstwie als natürliches Phänomen reproduzierbar. Gentechnologisch ist vielleicht in Zukunft einiges machbar, nur wird man diesen Bereich intelligent gezielten Gentransfers unter systematischer Umgehung sämtlicher natürlicher Schranken wohl kaum als natürliches Phänomen bezeichnen wollen. Wenn sich das als einzige Methode systematischen Aufbaus neuer genetischer Information in den Spezies erweist - und alles spricht bislang dafür - dann beweist diese Methode zugleich die Notwendigkeit intelligenter Steuerung zur Bildung neuer Arten.
Weiteres zum Axiomensystem der gegenwärtigen Naturwissenschaft vgl. pp. 284-290. Die grundsätzliche und absolute und manchmal sogar feindselige Ablehnung der Frage nach dem Konstrukteur auch bei den komplexesten und genialsten Konstruktionen in der Natur ist für mich kein Axiom, sondern ein weltanschauliches Dogma.
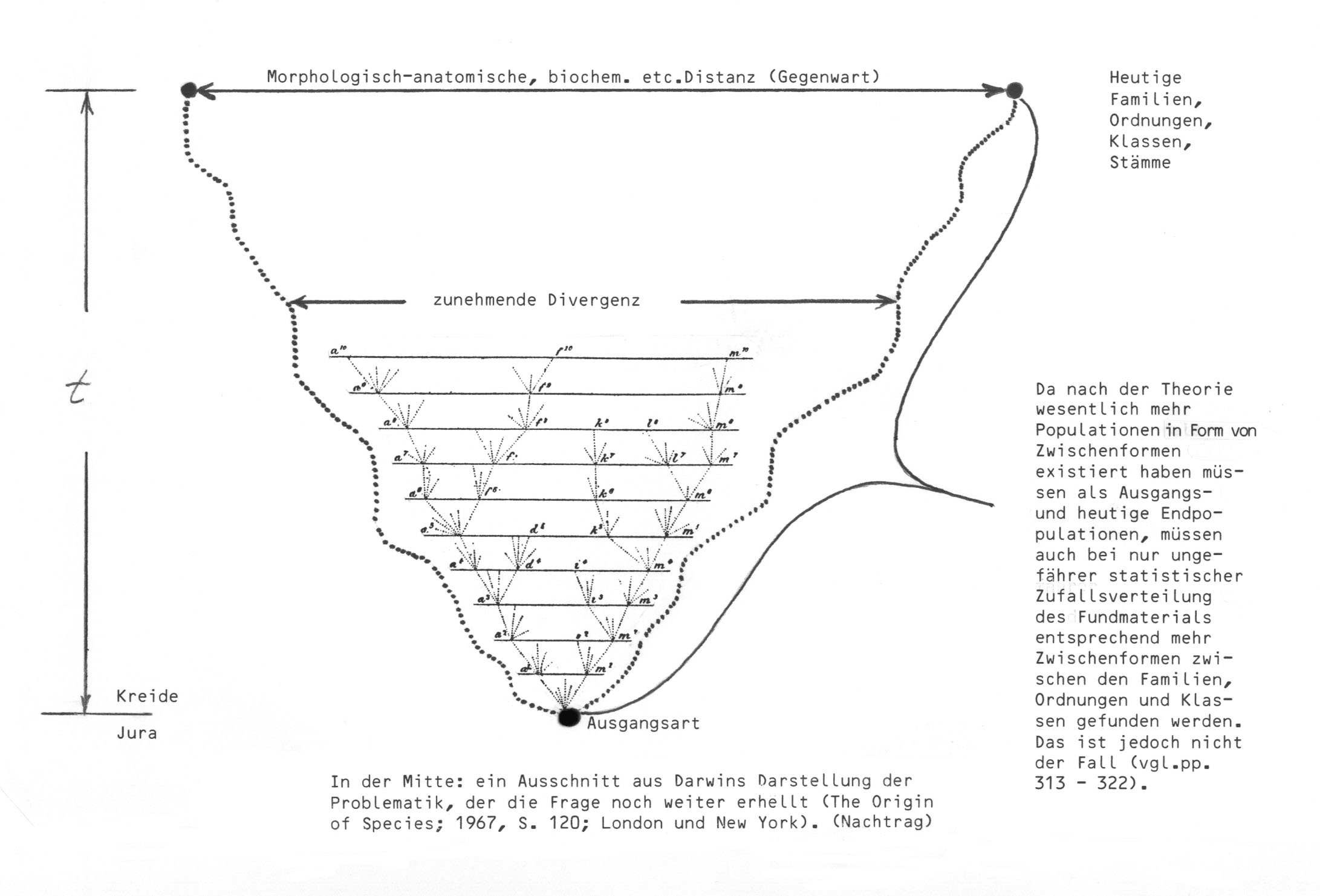
S. L.: "Sie fordern, dass zwischen verschiedenen Familien, Klassen etc. Übergangsformen existiert haben müssen. Hier ist zu bedenken: Was jetzt Familien und Klassen sind, waren nach der Evolutionstheorie vor langer Zeit nur Angehörige einer Art (oder einiger Arten, um Bastardierungen zu berücksichtigen) innerhalb einer Familie, die es damals gab. Die Unterschiede zwischen heutigen Familien etc. haben sich dann im Laufe der Zeit herausgebildet (schneller oder langsamer), und je weiter die Differenzierung gekommen ist, umso weniger ist das Vorkommen von Zwischenstufen denkbar, die entweder ein Mosaik oder intermediäre Stadien zwischen den Extremen aufweisen."
W.-E. L.: Vgl. dazu pp. 321 und 322, wo ich diesen Punkt detailliert behandelt habe.
So ein Stammbaum müsste dann ja ungefähr folgendermaßen aussehen:
Zum letzten Satz: Heute ja, aber nicht paläontologisch!
S. L.: "Ich finde es plausibel, dass Unterschiede im Genom mit der Zeit immer größer werden, nachdem sie sich bei der Rassenbildung einmal angefangen haben, herauszubilden."
W.-E. L.: Diese lineare Extrapolation ist als generelle Aussage sicher so nicht haltbar, wie uns das die lebenden Fossilien sehr nachdrücklich veranschaulichen. Auch zahlreiche Befunde aus der Pflanzen- und Tiergeographie widersprechen Ihrer Aussage (vgl. pp. 119/120, 412-419, speziell p. 418). Allenfalls für neutrale Punktmutationen (Kimura) ist eine lineare Extrapolation denkbar, aber selbst hier gibt es noch erhebliche Probleme (vgl. pp. 399 - 402 und 552 ff.).
Auf die Menschenrassen bezogen bin ich überzeugt, dass weder Mutation noch Isolation, noch Hybridisierung, noch Selektion zu neuen Menschen-Arten, -Gattungen, -Familien und -Ordnungen etc. führen werden. Subspecies non sunt species in statu nascendi!
Gilt der letztere Satz uneingeschränkt? Sicher nicht für die systematischen, neodarwinistischen und genetischen Artbegriffe (pp. 38 - 324). Aber eine Beobachtung ist immer wieder gemacht, ja als eine Gesetzmäßigkeit 1000mal festgestellt worden: Jede Variabilität läuft bei steter Selektion asymptotisch gegen ein Limit (vgl. pp. 351 - 372, speziell 357). Darüber hinaus ist die mit der Zeit zunehmende Unterschiedlichkeit von Genomen weitgehend mit Blockierung, Abbau und Verlust genetischen Materials gleichzusetzen (p. 438 ff. - siehe auch p. 473). Ich betone diesen Punkt hier immer wieder, weil heutzutage diese Tatsache weitgehend vergessen wird. Auf die Kategorien der biologischen Systematik angewandt, liegen diese Limits von Fall zu Fall verschieden: in manchen Fällen bleiben sie innerhalb der Spezies, in anderen gehen sie bis zur Gattung oder Familie und wohl sehr selten sind sie bei noch höheren Kategorien zu finden (siehe Ausführungen zum genetisch-plasmatischen Artbegriff).
Gehen wir mit der Schöpfungslehre von primären Arten mit ursprünglich maximaler genetischer Informationshöhe und größter individueller Anpassungsfähigkeit an die verschiedensten Umweltverhältnisse aus, so sind auch die meisten heutigen größeren Genomunterschiede bei verwandten Formen im Sinne von ARTBEGRIFF, p. 473, zu verstehen.
S. L.: "Das beobachten wir doch auf Schritt und Tritt: als Schüler lernen wir noch ähnliche Dinge, und je weiter der Lebensweg führt, desto größer werden die Unterschiede in den Kenntnissen, die der Einzelne besitzt."
W.-E. L.: Bedauerlicherweise sind Kenntnisse nicht vererbbar. Diese Art "geistiger Modifikation" lässt sich nicht für Evolutionsfragen einsetzen.
S. L.: "Sobald in tropischen Gewässern Hartsubstrat da ist, entwickeln sich auf ihm Korallen, und so ein Lebensraum wird dann zusehends verschieden von anderen, auf denen keine Korallen wachsen."
W.-E. L.: Es siedeln sich vorhandene Arten an, aber neue bilden sich dabei nicht.
S. L.: "Sie kritisieren, dass ich mich weigere, Kenntnislücken als Widersprüche zu einer Theorie zu akzeptieren. Logisch gesehen ist mein Argument völlig richtig, und folglich ist das Argument auch benutzbar."
W.-E. L.: Ich kritisiere selbstverständlich nicht, dass Sie sich weigern, Kenntnislücken als Widersprüche zu einer Theorie zu akzeptieren. Kenntnislücken gibt es in den Naturwissenschaften immer und an den verschiedensten Stellen und sind dazu da, durch die weitere Forschung zunehmend ausgefüllt zu werden. Was ich nicht verstehe, ist etwas ganz anderes, nämlich dass viele Neodarwinisten naturwissenschaftliche Argumente und Tatsachen, die ihrer Theorie widersprechen, grundsätzlich in die Kategorie der Kenntnislücken zur Rettung der Theorie einordnen. "Kenntnislücke" soll ja immer heißen: Wir wissen nur im Moment noch nicht wie ein bestimmtes Phänomen, etwa der Fangapparat von Utricularia vulgaris, mit der Theorie zu erklären ist, aber wenn wir nur an der Theorie festhalten, wird sie uns das schon eines Tages noch mit Mikromutationen, Rekombination, Selektion etc. erklären.
Es könnte sich ja auch um eine prinzipielle Lücke in der Struktur der Theorie handeln, so dass bestimmte Phänomene wie die Synorganisationen von Utricularia bis zum Auge und Gehirn niemals mit der Theorie erklärt werden können. Und dass diese Möglichkeit angesichts der ungeheuer komplizierten Konstruktionen in der Natur nicht ernsthaft in Erwägung gezogen wird, das ist es, was ich beanstande. Und genau an dieser Stelle erhebt sich wieder die Frage nach den Falsifikationskriterien der Theorie.
Grundsätzlich alle mit einer Theorie nicht erklärbaren Einwände, Argumente und Tatsachen als Kenntnislücken einzustufen, bedeutet ja eine Totalimmunisierung einer solchen Theorie, - die damit zum unwissenschaftlichen Dogma wird. Denn sämtliche falschen Behauptungen, Hypothesen und Theorien aller Wissenschaftszweige könnte man mit dieser Methode gegen jegliche Widerlegung immunisieren, - was ja wohl nicht der Sinn einer wissenschaftlichen Erklärung sein kann.
S. L.: "Richtig finde ich den von Ihnen aufgegriffenen Gedanken, dass zwei Frösche, die die einzigen Überlebenden von 120.000 befruchteten Eiern sind, wohl kaum als die Besten angesehen werden können, dass vielmehr der Zufall in der Selektion eine große Rolle gespielt haben muss. Diese Fälle werden allgemein zu wenig berücksichtigt von Populationsgenetikern. Im Froschbeispiel spielt der Zufall in der Tat eine große Rolle, doch halte ich die evolutionsbiologische Schlussfolgerung für zulässig, dass die Selektion hier dazu geführt hat, dass die Frösche eben wegen der hohen Verlustrate 120.000 und nicht nur 20 Eier erzeugen."
W.-E. L.: Mich wundert bei solchen Beispielen nur immer, warum es da keine Selektion gegen "Verschwendung" gibt.
S. L.: "Bei der Geburtshelferkröte werden erheblich weniger Eier erzeugt, weil wegen der Brutpflege noch immer genug Junge überleben."
W.-E. L.: Das erinnert mich an eine Verwandte der Geburtshelferkröte, die Wabenkröte als schwieriges Beispiel für die Synthetische Evolutionstheorie. Wie soll man sich ihre Brutpflege durch kaum sichtbare Mikromutationen und Selektion über große Zeiträume von Hunderttausenden bis Millionen von Jahren in Tausenden von kleinen Schritten vorstellen? Aber selbst wenn man sich einen solchen Prozess mit viel Phantasie noch ausmalen kann, wie wollte man ihn naturwissenschaftlich empirisch beweisen? Und schließlich erhebt sich wieder die Frage nach den Falsifikationskriterien einer solchen Theorie.
S. L.: "Wenn Evolutionsbiologen sagen, die Evolution werde durch genetische Variation und Selektion vorangetrieben, so stimme ich auch nicht zu."
W.-E. L.: Erfreut stelle ich fest, dass Sie sich damit von der Auffassung der meisten Befürworter der Synthetischen Evolutionstheorie ganz klar distanzieren (vgl. dazu die Dokumentation pp. 570 - 583)
S. L.: "Das Prinzip von Variation und Selektion ist vielfältig in der Ontogenese und Phylogenese wirksam, was man berücksichtigen sollte. Außerdem gibt es viele Variationen von allem Möglichen, die den Verlauf der Evolution beeinflussen, wobei genetische Variationen, eben nur die genetische Information variieren und nicht mehr."
W.-E. L.: Die genetische Variation ist jedoch nach der Synthetischen Evolutionstheorie die einzige, die vererbt wird und damit auch die einzige, die den Aufbau neuer genetischer Information und damit den Aufbau aller Strukturen in der Phylogenese besorgen soll. Die Variation von allem Möglichen und die Bedeutung der nichtgenetischen Faktoren bei der Umsetzung der genetischen Information betone ich hier, weil diese Faktoren die genetischen Mikroveränderungen bei weitem überlagern können und somit als "Störfaktoren" ein weiteres Problem für die Theorie sind.
S. L.: "Insgesamt lerne ich recht viel aus dem Studium kritische Schriften zur Evolutionstheorie, so auch aus der Ihrigen. Dennoch stehe ich nicht nur zeitgenössischen Darstellungen zur Evolutionstheorie, sondern auch den kritischen Gegenschriften kritisch gegenüber, denn in den letzteren findet man sehr wohl hanebüchene Argumentationsfehler."
W.-E. L.: Hier haben wir wieder vieles gemeinsam. Ich betrachte zeitgenössische Darstellungen zur Schöpfungslehre genauso kritisch wie die der Evolutionstheorie. In beiden habe ich schon "hanebüchene Argumentationsfehler" gefunden. Doch habe ich auch so manche Anregung erhalten.
S. L.: "Vieles an evolutionstheoretischen Vorstellungen ist noch unausgegoren, doch vieles halte ich für gut begründet und plausibel, so dass ich für mich keinen zwingenden Grund sehe, von der Evolutionstheorie abzurücken."
W.-E. L.: Aufgrund der komplexen Synorganisationen, der Ergebnisse der Mutationsforschung und der paläontologischen Situation hat sich bei mir die Überzeugung gefestigt, dass weite Bereiche der biologischen Realitäten nicht mit der Synthetischen Evolutionstheorie vereinbar sind. Alle Erfahrung lehrt vielmehr, dass bereits die nicht annähernd komplexen kybernetischen Systeme der menschlichen Technik [immer] den intelligenten Konstrukteur erfordern. Wieviel mehr trifft das erst auf die tausendmal besseren kybernetischen Konstruktionen in der Natur zu!
S. L.: "Noch eine kleine Bemerkung zum Äußeren Ihres Buches: Die Schrift ist stellenweise derart winzig, dass es nur noch mit detektivischer Mühe möglich ist, den Text zu lesen. Sie sollten bei einer neuen Auflage sämtliche winzigen Schriftsätze durch lesbare erneuern. Wie wäre es, wenn Sie überhaupt einen Schreibcomputer benutzen würden? Ich selbst will anfangen, einen solchen zu benutzen. Für dickere Arbeiten ist er geradezu ideal."
W.-E. L.: Sie haben völlig Recht. Die erste Reaktion von Herrn Dr. Stüber, Computerspezialist hier (in Köln) am MPI, auf meine Arbeit war, mich mit den Spitzenleistungen der neuesten Technik bekannt zu machen, und ich kann diese Leistungen nur hoch einschätzen. Mein Problem ist jedoch, dass ich in nächster Zeit durch vielfältige Aufgaben in der praktischen Forschung nicht dazu kommen werde, die inzwischen bald 600 Seiten in den Computer zu geben. Aber vielleicht ergibt sich demnächst doch noch eine vernünftige Alternative. Herzlichen Dank jedenfalls für die Anregung!
S. L.: (P.S.) "Noch eine weitere Bemerkung: Von Evolutionskritikern, so auch von Ihnen, wird immer wieder gesagt, die bisherigen Kenntnisse gestatten es nicht, den Übergang von einer zu einer ganz anderen Struktur zu erklären, weil intermediäre Stadien offenbar nicht funktionsfähig seien. Die Synergetik, wie Hermann Haken sie betreibt, hat mittlerweile gezeigt, dass es auch in der Physik plötzliche Übergänge von einem zu einem anderen Ordnungszustand gibt, ohne dass es beliebig viele Übergänge gibt. Beim Laserlicht springen Elektronen im Gleichtakt auf eine nächst niedrige Umlaufbahn und geben hierbei einen Lichtquant ab. Dies geschieht aber erst bei hinreichend hoher Energiezufuhr, vorher sind die Elektronensprünge nicht im Gleichtakt, und es ist lediglich die Gesamtzahl der Elektronensprünge proportional zur Energiezufuhr. Einen Übergang zwischen dem normalen und dem Laserlicht gibt es nicht. Solche Qualitätssprünge sind typisch für alle Systeme, die weit ab sind vom thermodynamischen Gleichgewicht. Da auch Lebewesen weitab vom thermodynamischen Gleichgewicht sind, erwarte ich derartige Qualitätssprünge auch bei ihnen, auch bei deren Evolution. In der Ontogenese gibt es genug von ihnen. Im Rahmen eines Axiomensystems, das ohne höhere Gewalt auskommt, lassen sich also Qualitätssprünge durchaus verstehen. Solange mein naturwissenschaftliches Axiomensystem nicht zu Widersprüchen führt, sehe ich keinen Anlass, es aufzugeben."
W.-E. L.: Im Gegensatz zu p. 446 stelle ich erneut fest, dass Sie Alternativen zur kontinuierlichen Evolutionstheorie durchaus aufgeschlossen sind. Ich kenne jedoch bisher weder empirische noch theoretische Belege dafur, dass etwa der Wechsel physikalischer Aggregatzustände oder physikalische Qualitätssprünge überhaupt, die Neubildung ganzer Serien spezifisch-informationsreicher DNA-Sequenzen, ganzer synorganisierter Gensysteme und deren ebenfalls spezifisch raumzeitliche Regulation und Expression in der Ontogenese verständlich machen könnten. Um wieder mein Paradebeispiel zu nehmen: Wie sollten physikalische Qualitätsprünge weitab vom thermodynamischen Gleichgewicht aus einem normalen Blatt die Kastenfalle von Utricularia produzieren? Hakens Erklärungsmodus erinnert mich an Conklins Bemerkung zur Zufallsentstehung des Lebens: "The probability of life originating from accident is comparable to the probability of the unabridged dictionary resulting from an explosion [ - Auch ein Vorgang weit ab vom thermodynamischen Gleichgewicht!] in a printing shop." Alle Erfahrung lehrt mich hingegen, dass der Ursprung kodierter komplexer Information immer eine geistige Ursache hat und nicht auf Zufälle zurückzuführen ist. Und das heißt ja nun keinesfalls die Aufgabe des ganzen naturwissenschaftlichen Axiomensystems, vielmehr die Erweiterung um die geistige Dimension für die Ursprungsfrage. Die Begründer der modernen Naturwissenschaften wie Kepler, Newton, Linné, Cuvier, von Baer, Mendel und viele andere sahen darin ja eine feste Basis und tiefste Sinngebung ihrer Arbeit, die zum naturwissenschaftlichen Axiomensystem generell erst führte. "...few scientists doubt, that there is something 'out there' to be examined...That pragmatic belief today is successor to a deeply held Christian conviction about the created world that attended the rise of modern science" (C.A. Russel 1988: Objectivity in science; Nature 331, 558). - Was zugleich eine Absage an den philosophischen Idealismus bedeutet. (Im übrigen besteht zwischen Christenheit und Bibel eine große Kluft.) Aber auf all diese verschiedenen Fragen möchte ich an dieser Stelle nicht weiter eingehen.
Abschließend möchte ich Ihnen noch einmal für Ihren Brief danken, und ich würde mich selbstverständlich auf weitere Kommentare von Ihnen freuen [Bisher (12 Monate später) noch nichts gekommen. Nach meiner etwa 20-jährigen schriftlichen Diskussionserfahrung zum Evolutionsthema endete die Diskussion regelmäßig mit meiner Antwort. Aber vielleicht kommen noch Kommentare.]
Mit freundlichen Grüßen,
W.- E. Lönnig
E. Trumler (1981, pp. 201/202) zur Bildung der Hunderassen durch Mutationen:
Mutationen sind - gemessen an den Wildformen - immer Fehlleistungen der Natur. Wenn zum Beispiel die Haare länger wachsen als bei den Wölfen üblich, dann ist jenes Gen in seiner Wirkungsweise behindert, das die Haarlänge kontrolliert. Wenn Haarlosigkeit auftritt, muss man folgern, dass genetisch die Fähigkeit zur Ausbildung des normalen Haarkleides verloren gegangen ist, und nicht anders ist es da, wo die Fähigkeit fehlt, unter dem Deckhaar eine Unterwolle mit jahreszeitlichem Rhytmus auszubilden. Alle Mutationen die wir bis heute kennen, sind nichts anderes als Fehlleistungen der Natur, wenn wir dabei die ursprüngliche Wildform des veränderten Haustiers als Maßstab nehmen. (Meine wilden Freunde. Die Wildhundarten der Welt. München.)
Trumler führt weiter aus, inwiefern diese 'Fehlmutationen' z.B. beim Münsterländer für den Menschen nützlich sind, aber bei keinem Wolf, Schakal oder Koyoten von Vorteil wären. Hinzufügen möchte ich, dass man sich durch Isolation (abgelegene Land- und Inselgebiete) das Überleben einiger Rassen in der Natur vorstellen könnte - wie das wohl beim Dingo der Fall ist - , aber in Konkurrenz mit den Wildtypen hätten sie wohl keine Chance. Bei der Entstehung der Wildhundarten selbst möchte ich an ökologisch relevante Alternativprogramme bei der Ausgangsform zur maximalen Umweltbewältigung denken. Aber es stehen auch neutrale Mutationen sowie Informationsverluste bis zum Niveau der heutigen Formen zur Debatte.