(5) HYBRID DYSGENESIS
Bevor wir auf die Bedeutung der Chromosomenmutationen für den Artbegriff zu sprechen kommen, wollen wir uns mit dem Phänomen der hybrid dysgenesis beschäftigen, einem Syndrom assoziierter, aberranter Merkmale bei Drosophila melanogaster, das unter diesem Namen erstmals von Kidwell, Kidwell und Sved 1977 beschrieben wurde und das unter anderem Sterilität verursacht. Obwohl der größte Teil der Syndrom-Kennzeichen schon wiederholt vorher beschrieben worden war, sind die Ursachen des Phänomens erst in den letzten Jahren genauer bekannt geworden. Wie sehr häufig bei neueren faszinierenden Entdeckungen mit noch unvollständig bekannter Tragweite, sind auch in diesem Falle von vielen Forschern große evolutionistische Erwartungen geäußert worden.
Ronsseray, Anxolabéhère und Périquet beschreiben in der Einleitung ihres Artikels HYBRID DYGENESIS IN Drosophila melanogaster: INFLUENCE OF TEMPERATURE ON CYTOTYPE DETERMINATION IN THE P-M SYSTEM (1984, p. 17) das Phänomen wie folgt:
The phenomenon of hybrid dysgenesis in Drosophila melanogaster (Kidwell et at. 1977) occurs in certain inter-strain crosses and groups a set of germline abnormalities including temperature-dependent sterility, high rates of mutation, male recombination, chromosome breakage and several other traits. Two independent systems of hybrid dysgenesis, called I-R and P-M, have been described (for review see Kidwell 1979; Sved 1979; Bregliano et al. 1980; Kidwell 1982) and are now known to be associated with two families of transposable elements: the I elements and the P elements (Bregliano and Kidwell 1983).
In the P-M system temperature-dependent sterility (gonadal dysgenesis) occurs when mates of a P strain (paternally contributing) are mated with females of an M strain (maternally cortributing) * but not in the reciprocal cross of intrastrain crosses. A third category has also been described, a so-called Q strain that gives nondysgenic progeny, at least for gonadal sterility, whatever way crosses with P or M strains are made (Engels 1979a; Kidwell and Novy 1979).
P and Q strains carry 30 - 50 copies of the P element family dispersed over all major Chromosome arms; in contrast old M laboratory strains do not generally carry P elements but the natural M populations exhibit copies of P elements (Kidwell 1983); Anxolabéhère et al., in preparation). The P elements isolated from hybrid dysgenesis-induced mutations are heterogenous in size (Bingham et al. 1982; Rubin et al. 1982); the largest element presently known is 2.9 kb long and its structure has been described by O'Hare ard Rubin (1983).
Although female offspring from a dysgenic cross M (female) x P (male) have the same chromosomal composition as the female offspring from the reciprocal cross, only the former may give, at 28,5° C, many dysgenetic progeny when mated to a P male. These females have inherited from their M female parent the extrachromosomal background, called M cytotype, which allows the transposition of P elements. Similarly, the daughters from P (female) x M (male) crosses have maternally inherited P cytotype and this extrachromosomal background is opposed to the P element transposition (Engels 1979b). However, cytotype switch is only partially independent of chromosomal determinants and is correlated with the number of P or M derived chromosomes from P or M strains (Kidwell 1981).
* "The phenomenology of hybrid dysgenesis is observed when males that carry P transposons are crossed with P- females". - Syvanen 1984, p. 284.
Sieben Jahre zuvor wiesen Kidwell et al. in ihrer Arbeit darauf hin, dass sechs der Charakteristika (erhöhte Mutationsrate, Sterilität, männliche Rekombination, Chromosomen-'transmission ratio distortion', chromosomale Aberrationen und örtliche Zunahme weiblicher Rekombination) schon vorher beschrieben wurden und dass sie selbst zusätzlich das erste Mal von Non-disjunction in diesem Zusammenhang berichten können.
Nach den obigen Ausführungen zum Artbegriff der Synthetischen Evolutionstheorie dürfte dem Leser sofort klar sein, warum mit diesem Syndrom starke entwicklungstheoretische Erwartungen verbunden worden sind: Sterilität und hohe Mutationsraten bilden die idealen Anknüpfungspunkte für neodarwinistische Erwägungen. Nach Hinweis auf die männliche Rekombination als dem ersten Punkt ihrer Beschreibung der 'dysgenic traits' schreiben Kidwell et al. in ihrer Originalarbeit (1977, pp. 816/817) zu den beiden Punkten Sterilität und Mutation:
Sterility: Sterility is defined as the complete absence of progeny in an individual after the opportunity to mate with at least two members of the opposite sex from a fully fertile strain. High frequencies of both male and female sterility have been found in some inter-strain crosses (Kidwell and Kidwell 1975a), though in contradistinction to Haldane's (1922) rule, female sterility tends to exceed male sterility. Dysgnic F1 females have been observed to accept and mate with males of both parental strains, although there were significant differences in mating speed between dysgenic and nondysgenic females of the same genotype (i.e., females arising from reciprocal crosses). In some crosses females produced eggs, while in others there was a complete absence of egglaying. Additional data on F1 A female sterility are provided in later sections of this paper.
Mutation: In both the first and second chromosomes of hybrids (Type A in our classification) Slatko and Hiraizumi (1973) observed lethal mutation rates up to ten times above normal, associated with the presence of second chromosomes from the Texas T-007 strain. Kidwell, Kidwell an Ives (1977) have also reported a 5-10 fold increase in X-linked lethal mutations in hybrids involving male parents from both the Cranston and South Amherst populations. These experiments indicate that the strain source of the monitored X chromosome was not critical to the production of high mutation rates. There was no evidence for nonrandom location of induced lethals within the X chromosome.
Zum Thema Sterilität möchten wir zunächst festhalten, dass die betroffenen Linien (trotz großer evolutionistischer Erwartungen) vernünftigerweise von keinem der damit befassten Autoren in den Rang eigener Arten erhoben worden sind. Vernünftigerweise - weil andernfalls der durch die Sterilität definierte Artbegriff nicht nur verschiedene Linien zu eigenen Arten erklären würde, sondern die Artgrenzen häufig durch sonst identische Genotypen gezogen werden müssten (wie wir das in ähnlicher Weise schon beim Reis und Weizen kennengelernt haben). Nach dem undifferenzierten Arttrennungskriterium der Sterilität als postzygotischem Isolationsmechanismus müssten jedoch zumindest alle F1-Weibchen, bei denen nach Paarung "a complete absence of egglaying" festgestellt wurde, zu einer anderen Drosophila-Art gehören. Statt die erhofften Evolutionserwartungen zu erfüllen, relativiert die Tatsache, dass aus überzeugenden Gründen kein Forscher hier neue Spezies aufstellt, die neodarwinistischen Artabgrenzungskriterien aufs Neue. In den vorliegenden Beispielen reicht weder die stark temperaturabhängige Sterilität beim P-M System noch die herabgesetzte Fertilität beim I-R System aus, um verschiedene Linien als eigene Arten zu führen. Bei der temperaturabhängigen Sterilität würde überdies der Artbegriff neben all den anderen bereits geschilderten reversiblen Umständen auch noch temperaturabhängig werden.
Engels schreibt 1983, p.324:
GD (Gonadal Dysgenic Sterility) is highly temperature dependent. In dysgenic females it reaches its highest level at 29° C and is essentially absent at 24° C. The frequency of sterility increases approximately linearly through the intermediate temperature. For a given P strain genome, GD sterility in males is normally only about one-forth as frequent as female sterility and requires higher temperatures for its expression.
Kidwell und Novy geben (1979, p. 1136) 24° bis 26° C für die Weibchen und 27° bis 29° C für die Männchen als 'change from permissive to restrictive temperature' an. Ab 24° C gehörten nach strikter Anwendung der neodarwinistischen Auffassung die Ausgangslinien der F1-Weibchen und ab 27° C die der Männchen zu einer anderen Art, - unterhalb dieser Temperaturen gehörten jedoch alle zur gleichen Art (- welche Artunterscheidung wohl niemand ernsthaft vertreten möchte).
Die von vielen Forschern wie Kidwell, Engels, Syvanen et al. mit dem Phänomen der Hybrid Dysgenesis verknüpfte evolutionistische Bedeutung ist keine induktive Schlussfolgerung, etwa derart, dass tatsächlich in irgendeiner Weise die Bildung neuer Spezies stattgefunden hätte (von höheren Kategorien wie Genera, Familien, Ordnungen und Klassen ganz zu schweigen), sondern eine Deduktion nach neodarwinistischen Voraussetzungen, nach denen Sterilität und Mutationen für den Ursprung und die Komplexität aller Lebensformen auf unserer Erde zusammen mit der Selektion die Hauptursachen bilden.
Mit der Widerlegung der Voraussetzungen fallen auch die evolutionistischen Erwartungen. Für den Genfluss habe ich das oben schon gezeigt, und für die von Slatko und Hiraizumi festgestellte (und von Kidwell et al. bestätigte) Verzehnfachung der Letalmutationsrate bei Anwesenheit des zweiten Chromosoms der Texas T-007 Linie (sowie für die beim I-R System festgestellten Deletionen - Picard 1978) sei zunächst hervorgehoben, dass Letalmutationen kein erfolgversprechender Ansatz für die Entstehung der Arten und höheren Kategorien sein können. Aber die Mutationsfrage umfasst hier noch eine ganze Reihe weiterer Aspekte, auf die wir auf den Seiten 325-463 zurückkommen möchten.
Zu dem durch die P-Elemente verursachten P-M System (vgl. Ronsseray et al. oben) sind einige neuere Entdeckungen hervorzuheben, die von Syvanen (1984, p. 285) wie folgt beschrieben worden sind:
P- flies are found in nature infrequently today, but they are common in laboratory strains taken from nature thirty years ago. In fact, P elements are not seen in flies isolated before 1945, making it appear that the P element has recently been introduced into wild-fly populations, as the "recent-invasion" hypothesis suggests (51). The P element could have originated either from an unrelated species and been introduced into Drosophila by means of viral transmission (70) or, less likely, could have arisen in an isolated Drosophila sub-population.
Alternatively, the "stochastic-loss" hypothesis proposes that all wild flies originally carried P elements but that the laboratory strains lost them (33). The major difficulty with this explanation is that the early flies are not just P- in their phenotype; they have no DNA sequences that cross hybridize to P element sequences. It is hard to imagine how the multiple copies could have become so completely deleted in such a short period of time. If this is true, we will also have to explain why P DNA is so much less stable than the other middle repetitive sequences in laboratory stocks. In any case, this hypothesis is testable by monitoring the long-term stability of P sequences in current laboratory stocks.
If we grant that the recent-invasion hypothesis is true, it means we have witnessed a major genetic change in fly populations that represents a substantial step toward a speciation event.
Abgesehen von der letzteren Behauptung, die in verschiedener Formulierung durch den ganzen Artikel geht und die wieder den neodarwinistischen Ansatz voraussetzt, ist bei Zutreffen der 'recent-invasion'-Hypothese das P-M System bei Drosophila melanogaster kaum mehr als 40 Jahre alt und kann damit schlecht den Ursprung dieser Art vor den postulierten Jahrmillionen erklären (im übrigen hat sich Drosophila seit ihrem Nachweis in dem auf 35 000 000 Jahren datierten miozänen Bernstein nicht wesentlich verändert). - Der I+ Phänotyp (bezogen auf das F1-Phänomen) machte 'a significant appearance in flies in the 1930s' und scheint damit nicht sehr viel früher als die P-Transposons bei Drosophila aufzutreten. Eine Hypothese macht für das Auftreten der I Transposons bei dieser Fliege den großräumigen Einsatz von DDT und für die P Invasion die Einführung von Organophosphat-Insektiziden verantwortlich und Syvanen meint u.a. (p. 287): "Perhaps it can be argued that some recent stress made Drosophila more vulnerable to I factor and P factor spread, without beneficial result to the fly."
Neben Syvanen haben viele weitere Autoren (z.B. Baltimore 1985, Emori et al. 1985, Paulson et al. 1985, Miller und Miller 1982, Temin 1980, Doolittle und Sapienza 1980, Orgel und Crick 1980 und viele andere) die Verwandtschaft bestimmter Gruppen von Transposons mit Retroviren hervorgehoben. So schreiben auch Finnegan, Hill, Bayer, Bowcock und Brown 1982, p. 33:
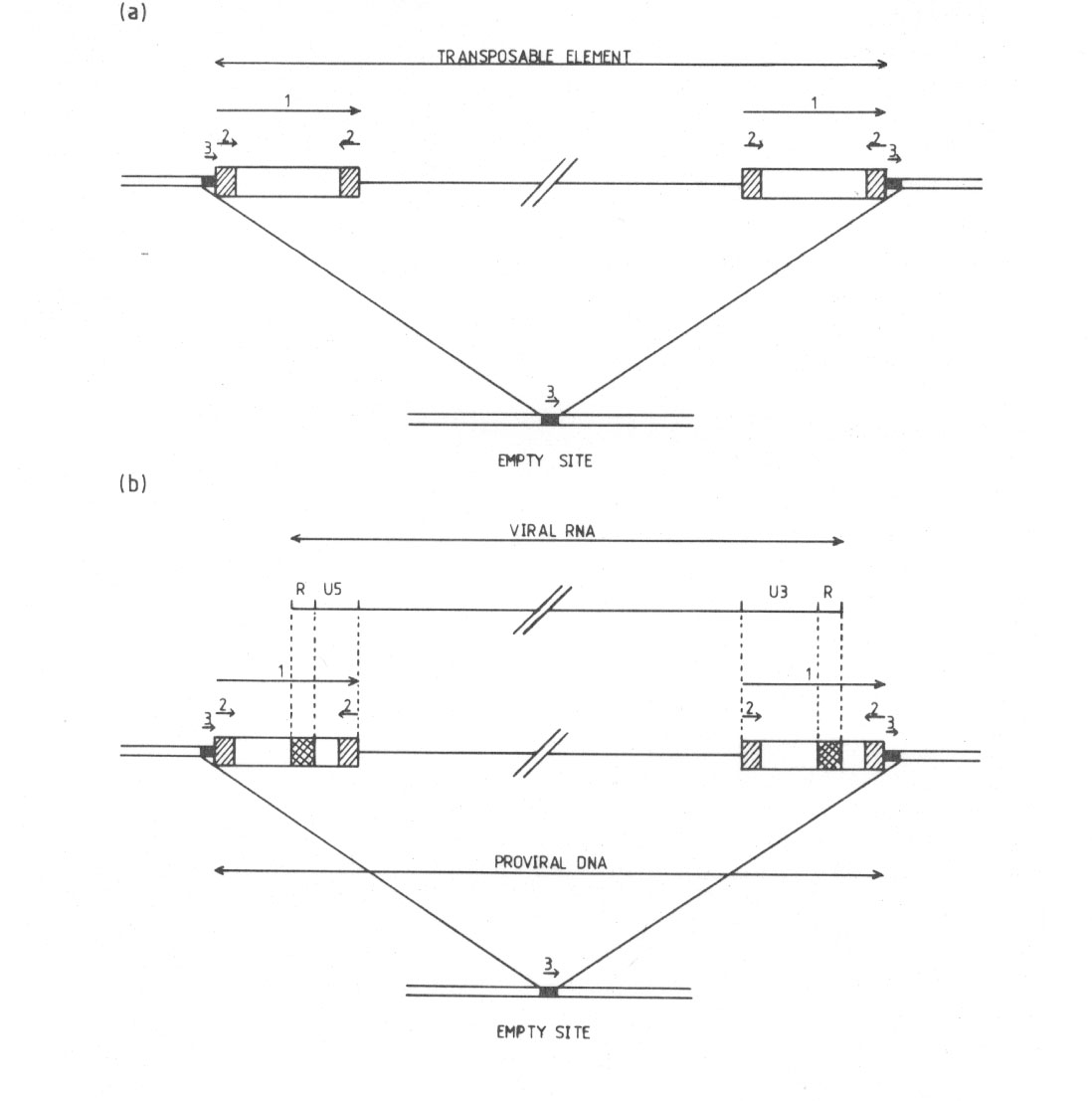
Several authors have noted the similarity between the sequence organization of transposable elements in Drosophila melanogaster and of the proviruses (Levis et al., 1980; Bayer et al., 1980). These two classes of elements (1) are of similar length, (2) are bounded by direct repeats a few hundred base-pairs long, which are themselves terminated by short inverted repeats, and (3) are flanked by a small number of bases, which occur once at the site of insertion in the absense of the element. The similarity extends to the first and last two base-pairs of each element. All proviruses that have been analysed start with the sequence T-G and end with C-A, and the same is true of copia, 412, and mdg 3 (but not 297) elements.
Abb. 24: (aus Finnegan et al. 1982): Strukturvergleich von Transposons mit Pro- und Retroviren.
FIG. 1 (a) Diagram of a typical eukaryotic transposable element, indicating the positions and orientations of the various repeat sequences. The relationship between a full and empty site is shown. The repeats are 1, long direct repeat; 2, short direct repeat; 3, repeat of a host sequence present once only at an empty site. (b) Diagram showing the relationships between an integrated provirus and a site into which it has inverted. U5, R und U3 are described in the text. Repeat sequences are described in (a) above.
Finegan et al. heben auf der Seite 34 ihrer Arbeit folgende weitere Punkte hervor.
One can easily imagine how proviruses could change their locations by going through a complete viral life-cycle (transcription of viral RNA, formation of extracellular virions, infection, reverse transcription, integration) or a partial life-cycle going directly from viral DNA and bypassing virions. The topological similarities between proviruses and transposable elements might indicate that they also move via a complete or partial life-cycle.
Mit der Retrovirenverwandtschaft wird verständlich, wie sich solche Elemente in relativ kurzer Zeit bei Drosophila und anderen Organismen verbreiten konnten.
Bei der Frage nach der Bedeutung der Transposons für die Entstehung der Arten und höheren systematischen Kategorien gehen die Meinungen weit auseinander - von hochgesteckten Erwartungen bis zur völligen Verneinung irgendwelcher evolutionistischer Ansatze. Ich möchte im Mutationskapitel auf das Thema zurückkommen (p. 438) und hier nur feststellen, dass bisher kein Nachweis für die Entstehung neuer Arten durch Transposons gegeben werden konnte, auch nicht im Sinne des morphologischen oder des neodarwinistischen Artbegriffs.
M. G. Kidwell stellt zu dieser Frage 1986, p. 170, fest:
Despite growing evidence for the widespread distribution of mobile elements in both prokaryotes and eukaryotes, their evolutionary significance remains obscure. Since the idea of "selfish DNA" was put forward by Doolittle and Sapienza (1980) and Orgel and Crick (1980), intense speculation has continued as to whether these elements are maintained and spread by virtue of their competitive edge in replication at the DNA sequence level despite a possible selective disadvantage or whether they are maintained, at least in part, by selection at a higher level of organization. However, very little is actually known about possible selective advantages or disadvantages that mobile elements might confer on individual organisms or supraorganismal groups over either the short or long term.
Ich halte es jedoch für möglich, dass durch die von Transposons verursachte Blockierung bzw. durch den Umbau genetischer Strukturen die weitere Forschung noch eine größere Bedeutung dieser Elemente für die Mikroevolution zutage fördern könnte, - was ebenfalls im Mutationskapitel weiter diskutiert werden soll.