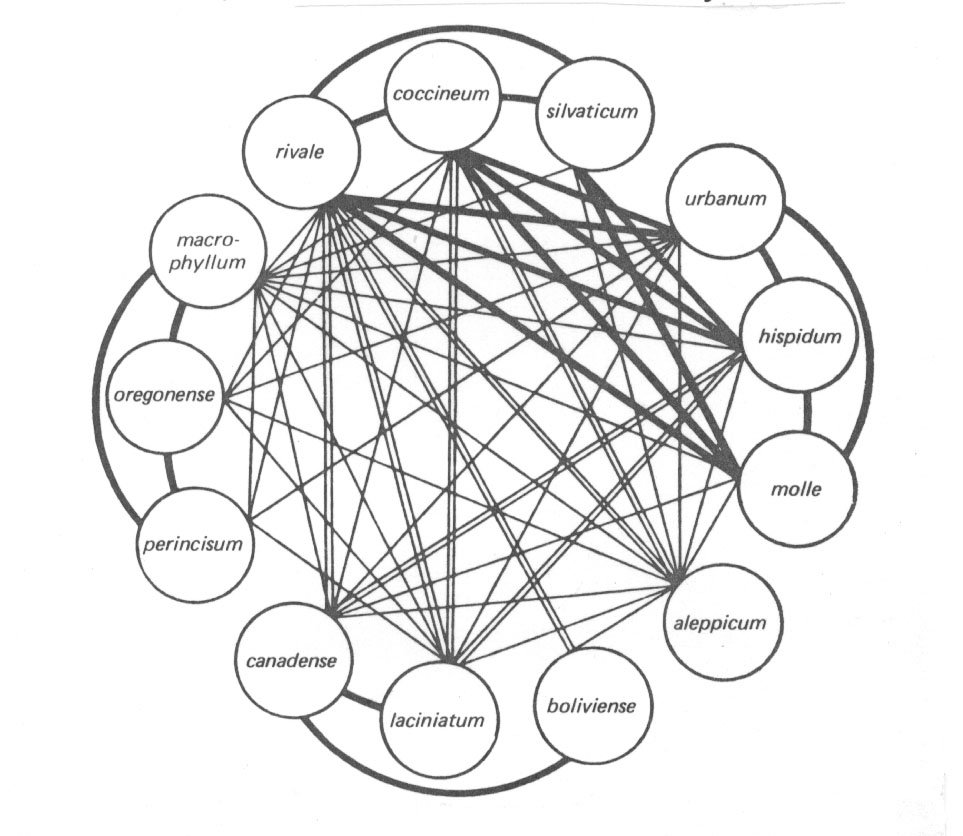
(7) KREUZUNGSPOLYGONE
Aus den bisher aufgeführten Tatsachen und Zusammenhängen erklären sich auch die oft sehr unterschiedlichen Fertilitätsgrade bei den Kreuzungspolygonen. Kommen wir noch einmal auf unser Geum-Beispiel zurück. Wie die folgende Abbildung (nach Gajewski aus Briggs und Walters 1984) zeigt, liegen auch bei dieser Rosaceengattung sehr unterschiedliche Fertilitätsverhältnisse vor:
Abb. 30: Fertility in F1 hybrids among different *Geum species. Thick lines indicate fertile hybrids; double lines partially fertile hybrids; thin lines sterile hybrids. Aus Briggs und Walters 1984.
*Grant führt (1981, p. 132) nach Gajewski u.a. noch folgende Punkte zu Geum auf: Das Genus umfasst etwa 56 Spezies, ist auf der Nordhalbkugel weit verbreitet und kommt auch in milderen Gebieten der südlichen Hemisphäre vor. Geum besteht (derzeit) nur aus Polyploiden von 4n bis 12n (n=7). Gajewsky unterteilt die Gattung in 11 Subgenera. Der größte Teil der Arbeit ist bisher in die Subgenera Oreogeum und Eugeum investiert worden. Letztere enthält 25 Spezies, von denen ein Teil in dem obigen Kreuzungspolygon autgeführt ist, u.a. unsere bekannten Arten Geum rivale und G. urbanum. "In the experimental garden the species can be intercrossed with ease. Any two species of the subgenus Eugeum can be crossed regardless of differences in ploidy level. And Eugeum can be crossed successfully with Oreogeum and with four other subgenera'' (Grant).
Es sieht fast so aus, als würden die Fertilitätsverhältnisse vieler 'Arten' untereinander den elementaren Grundsätzen der Logik widersprechen:
|
Die Hybriden sind: |
|||
|
Geum aleppicum |
x |
Geum coccineum |
partiell fertil |
|
G. coccineum |
x |
G. molle |
voll fertil |
|
G. molle |
x |
G. aleppicum |
steril |
|
Geum aleppicum |
x |
Geum rivale |
partiell fertil |
|
G. rivale |
x |
G. molle |
voll fertil |
|
G. molle |
x |
G. aleppicum |
steril |
|
Geum aleppicum |
x |
Geum rivale |
partiell fertil |
|
G. rivale |
x |
G. boliviense |
partiell fertil |
|
G. boliviense |
x |
G. aleppicum |
steril |
|
Geum aleppicum |
x |
Geum rivale |
partiell fertil |
|
G. rivale |
x |
G. urbanum |
voll fertil |
|
G. urbanum |
x |
G. aleppicum |
steril |
|
Geum aleppicum |
x |
Geum canadense |
steril |
|
G. canadense |
x |
G. rivale |
partiell fertil |
|
G. rivale |
x |
G. aleppicum |
partiell fertil |
|
Geum laciniatum |
x |
Geum coccineum |
partiell fertil |
|
G. coccineum |
x |
G. urbanum |
voll fertil |
|
G. urbanum |
x |
G. laciniatum |
steril |
|
Geum laciniatum |
x |
Geum canadense |
voll fertil |
|
G. canadense |
x |
G. coccineum |
steril |
|
G. coccineum |
x |
G. laciniatum |
partiell fertil |
|
Geum coccineum |
x |
Geum hispidum |
voll fertil |
|
G. hispidum |
x |
G. canadense |
partiell fertil |
|
G. canadense |
x |
G. coccineum |
steril |
Und so weiter. Unsere obigen Beispiele (F1-Sterilität bzw. -Letalität bei Reis-, Weizen- und Gerstensorten, der "Hybridensterblichkeit" beim Menschen sowie den Rekombinationsquadraten für epistatische Geninteraktionen) haben gezeigt, worauf solche Fertilitätsunterschiede genisch beruhen können.
(Soweit mir die Literatur bisher zugänglich ist, sind die Fragen nach den genauen genischen Ursachen für die hier aufgeführten Geum-Beispiele noch nicht geklärt.) Das Prinzip dürfte aber mit den obigen Ausführungen zur F1-Sterilität klar geworden sein: Partielle Fertilität der Hybriden aus Geum aleppicum x G. coccineum sowie volle Fertilität der Hybriden aus coccineum x molle sagen noch nichts über die Fertilität der Nachkommen von molle x aleppicum aus, weil in jedem Falle unterschiedliche Geninteraktionen mit spezifisch unterschiedlichen Auswirkungen auf die Fertilitätsverhältnisse möglich sind und offensichtlich auch in all den zitierten Geum-Beispielen real auftreten. Genische Differenzen (wie Modifikationen oder Ausfälle von Genfunktionen), die bei den Nachkommen aus der Kreuzung coccineum x molle noch mit Funktionsausgleich einhergehen (was z.B. auf die meisten rezessiven Mutationen zutrifft), führen bei molle x aleppicum zur Fertilitätsblockade durch Epistasie. Der Unterschied kann in solchen Fällen darauf beruhen, dass die Sterilität durch nicht mehr voll funktionsfähige Enzyme oder Hybridenzyme bzw. funktionsuntaugliche Genprodukte, die
Sand im übrigen hochempfindlichen physiologischen Getriebe darstellen, hervorgerufen wird. Qualitative Unterschiede in den Genprodukten (z.B. Variation in der räumlichen Struktur ursprünglich voll funktionsfähiger Enzyme) reichen dafür im Prinzip aus. (Die Aufführung solcher realistischer Möglichkeiten im Sinne der obigen Rekombinationsquadrate enthebt uns selbstverständlich nicht der Aufgabe, den Ursachen für den konkreten Fall genau nachzugehen.)
Es sei noch betont, dass solche (und stärkere) Sterilitätsbarrieren genauso zwischen Formen vorkommen, die nach herkömmlicher Systematik zu ein- und derselben Art gehören. Briggs und Walters führen (1984) dafür Layia glandulosa, Nigella arvensis und Elymus glaucus auf. Im Detail habe ich dafür oben schon das Reis- und das Weizenbeispiel zitiert.
Zu den Artkreuzungen bei der Gattung Mimulus stellen Briggs und Walters in Anlehnung an Vickery fest (p. 189):
Some interspecific hybrids proved relatively infertile, whilst other F1 hybrids were fully fertile. Some intraspecific crosses included in these experiment gave fully infertile hybrids, e.g. within M. laciniatus and M. nasutus. Paradoxically, some interspecific crosses between M. laciniatus and M. nasutus proved fully fertile.
Obwohl die Autoren meinen, dass Kreuzungsexperimente dieser Art eine wertvolle Informationsquelle zur Speziationsfrage sind, schließen sie ein Wort der Vorsicht zur Interpretation solcher Kreuzungsbefunde an, aus dem ich die Hauptpunkte zitieren möchte (p. 190):
First und foremost, only small numbers of plants are used in many crossing programmes and, in the extreme case, a single plant* is taken as a representative of a population! Crossing experiments are often halted at the F1 stage, as to raise F2 families a large amount of garden space may be needed. There are other grounds for caution. Glasshouses are frequently used in breeding experiments. Insect pests and fungal diseases may suddenly reach epidemic proportions during a series of crossing experiments, und success or failure of crossing may be influenced by such pests und diseases. The effects of bagging flowers must also be considered. Geiger (1965) reviews the relevant literature, which indicates that temperature inside pollen/insect-proof bags might be up to 15°C higher than ambient temperatures in the daytime and l-2°C lower at night. As pollen sterility und other effects may be induced at high temperatures, failure in crossing may be due to these external factors rather than to intrinsic differences. Furthermore, pollen 'fertilities' are commonly estimated, not by direct study of germinability or in a crossing test, but by staining with acetocarmine or other stain. Fully formed grains with nuclear staining are assessed as 'good' pollen, und mis-shapen, undersized, inadequately stained grains are judged to be 'bad'. The reliability of staining as an indicator of fertility is rarely, if ever, put to the test ... In assessing crosses, the possible complicating factor of genetic incompatibility is often forgotten.
*Bei meinen Untersuchungen an den Antirrhinum-Arten majus und siculum war nach umfangreichen Kreuzungen zwar ein guter Samenansatz festzustellen, aber es konnten nur 33 Pflanzen aus ca. 10 000 Samen (400 Kapseln) gezogen werden. Alle anderen Antirrhinum-Kreuzungen verliefen dagegen völlig normal (1986: Einhunderttausend F1-Pflanzen!). Der Anteil keimfähiger Samen aus der Kreuzung A. siculum X A. majus betrug damit nur 0,33 %. Die F1-Pflanzen selbst sind jedoch fertil. Mit nur einer oder wenigen Kreuzungen hätte man leicht auf eine absolute Barriere schließen können (Literatur zu früheren Untersuchungen bei Stubbe 1966).
Werfen wir einen Blick auf das Thema Selbststerilität:
Gottschalk weist (1984, p. 134) darauf hin, dass Selbstinkompatibilität (Selbststerilität) bei zahlreichen Pflanzen als artkonstantes Merkmal auftritt. "Sie ist bei einigen Kulturpflanzen von großer praktischer Bedeutung. Selbststerilität liegt vor, wenn
- die Selbstbestäubung einer Blüte,
- die Geschwisterbestäubung zwischen Blüten der gleichen Pflanze
- oder die Bestäubung von Blüten verschiedener Pflanzen des gleichen Genotypus
nicht zur Befruchtung führt. Diploide selbstinkompatible Pflanzen sind heterozygot für 2 verschiedene Allele des S-Locus, die einer multiplen Serie angehören. Eine Befruchtung ist nur dann möglich, wenn das S-Allel des Pollens nicht mit den Allelen im Griffel übereinstimmt." (Einzelheiten entnehme man Lehrbüchern der Genetik.)
Aufgrund solcher Gegebenheiten können Hybriden als "steril" klassifiziert werden, obwohl sie es bei genauerer Untersuchung - Feststellung von Selbststerilität - gar nicht sind.
Günther hebt (1984, p. 226) desweiteren hervor, dass die S-Allele auch die Kreuzbarkeit der 'Arten' zu kontrollieren scheinen. "Eine Reihe von Artkreuzungen ist in die Kreuzungsrichtung selbstinkompatibler Elter als Mutter X selbstfertiler Elter als Vater nicht möglich, wohl aber die reziproke Kreuzung."
Es dürfte klar sein, dass sich bei ausgeprägten chromosomalen Unterschieden weitere Schwierigkeiten bei Artkreuzungen ergeben können. Grant erwähnt (1981, p. 138), dass sowohl chromosomale als auch genische Faktoren bei Hybriden der Kompositengattung Layia (z.B. zwischen den Arten mit n = 7 und n = 8 Chromosomen) zu reduzierter Chromosomenpaarung führen. Bei Madia (die ebenfalls zu den Kompositen gehört), liegen ähnliche Verhältnisse vor (vgl. Abb. 31, nächste Seite). Für weitgehend chromosomal bedingte Inkompatibilitäts-Barrieren führt Grant folgende Genera auf (pp. 139/140):
|
Brassica (Cruciferae) |
(Yarnell 1956) |
|
Spergularia (Caryophyllaceae) |
(Ratter 1976) |
|
Atriplex triangularis group(Chenopodiaceae) |
(Gustafsson 1973) |
|
Nicotiana, annual sections(Solanaceae) |
(Avery 1938; Goodspeed 1954) |
|
Collinsia (Scrophulariaceae) |
(Garber and Gorsic 1956; Garber 1957, 1960) |
|
Microseris (Compositae) |
(Chambers 1955) |
|
Helianthus (Compositae) |
(Heiser, Martin, and Smith 1962) |
|
Stephanomeria (Compositae) |
(Gottlieb 1971) |
|
Bromus section Ceratochloa(Gramineae) |
(Stebbins and Tobgy 1944; Stebbins, Tobgy, and Harlan 1944; Hall 1955) |
|
Avena (Gramineae) |
(Rajhathy and Thomas 1974) |
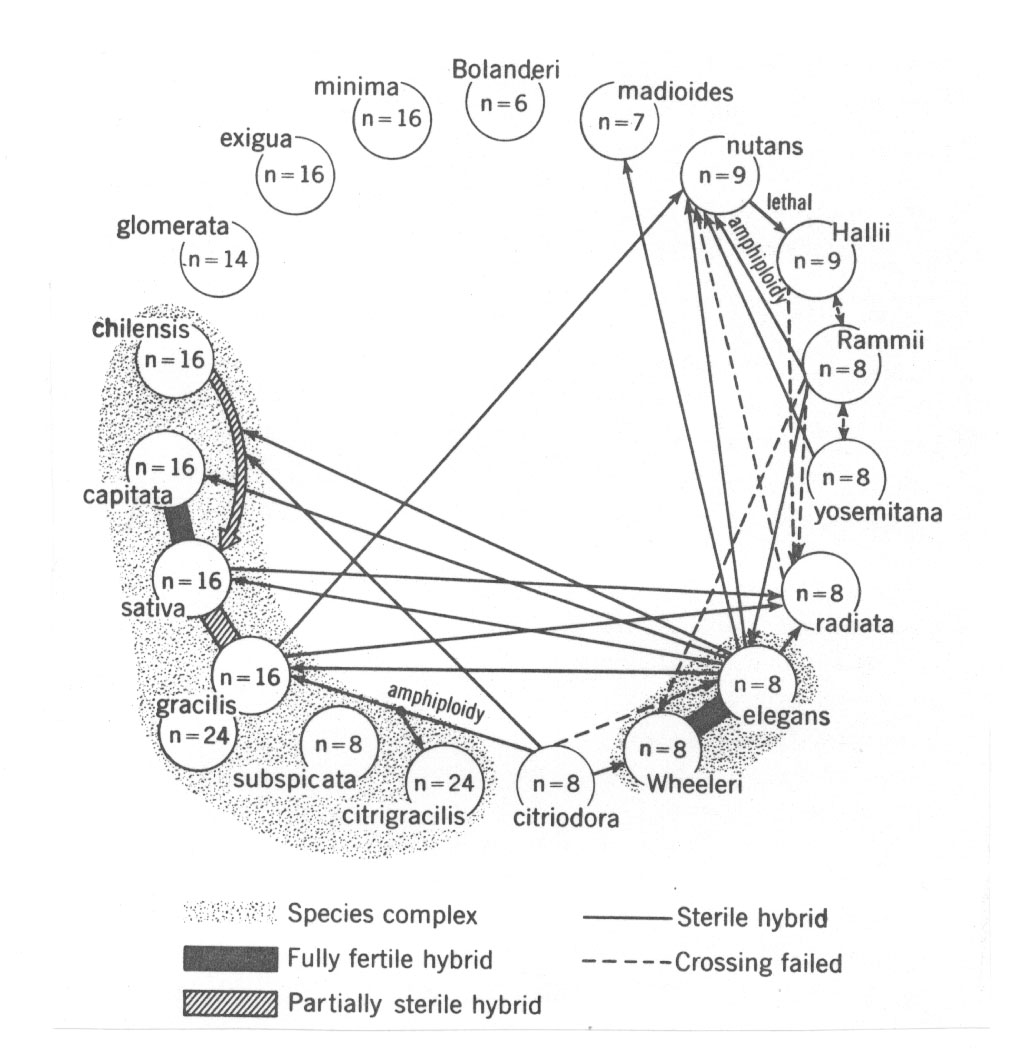
Abb. 31: Fertilitätsverhältnisse bei der Kompositengattung Madia (nach Grant 1981 aus Clausen). Die F1-Sterilität scheint hier häufig chromosomal bedingt zu sein.
Zur Frage nach den Ursachen der oft unterschiedlichen Fertilitätsverhältnisse innerhalb verschiedener Pflanzengenera sei noch eine vergleichende Untersuchung Grants erwähnt, nach welcher eine Korrelation der Lebensform mit den Fertilitätsverhältnissen festzustellen ist. Aufgrund seiner Literaturstudien über 72 taxogenetisch untersuchten Pflanzengruppen stellt er fest (pp. 143/145):
Species belonging to the same section or species group usually cross freely in woody and perennial herbs, but are usually separated by incompatibility barriers in annual herbs. The distribution of hybrid sterility barriers shows a similar correlation with life form F1 hybrids between related species are usually fertile or semifertile in woody plants and perennial herbs, but are mostly sterile in annual herbs.
Für den letzteren Punkt gibt es zwar zahlreiche Ausnahmen, dennoch scheint sich in der folgenden Tabelle eine klare Tendenz wiederzuspiegeln:
Tabelle 2 (aus Grant 1981):

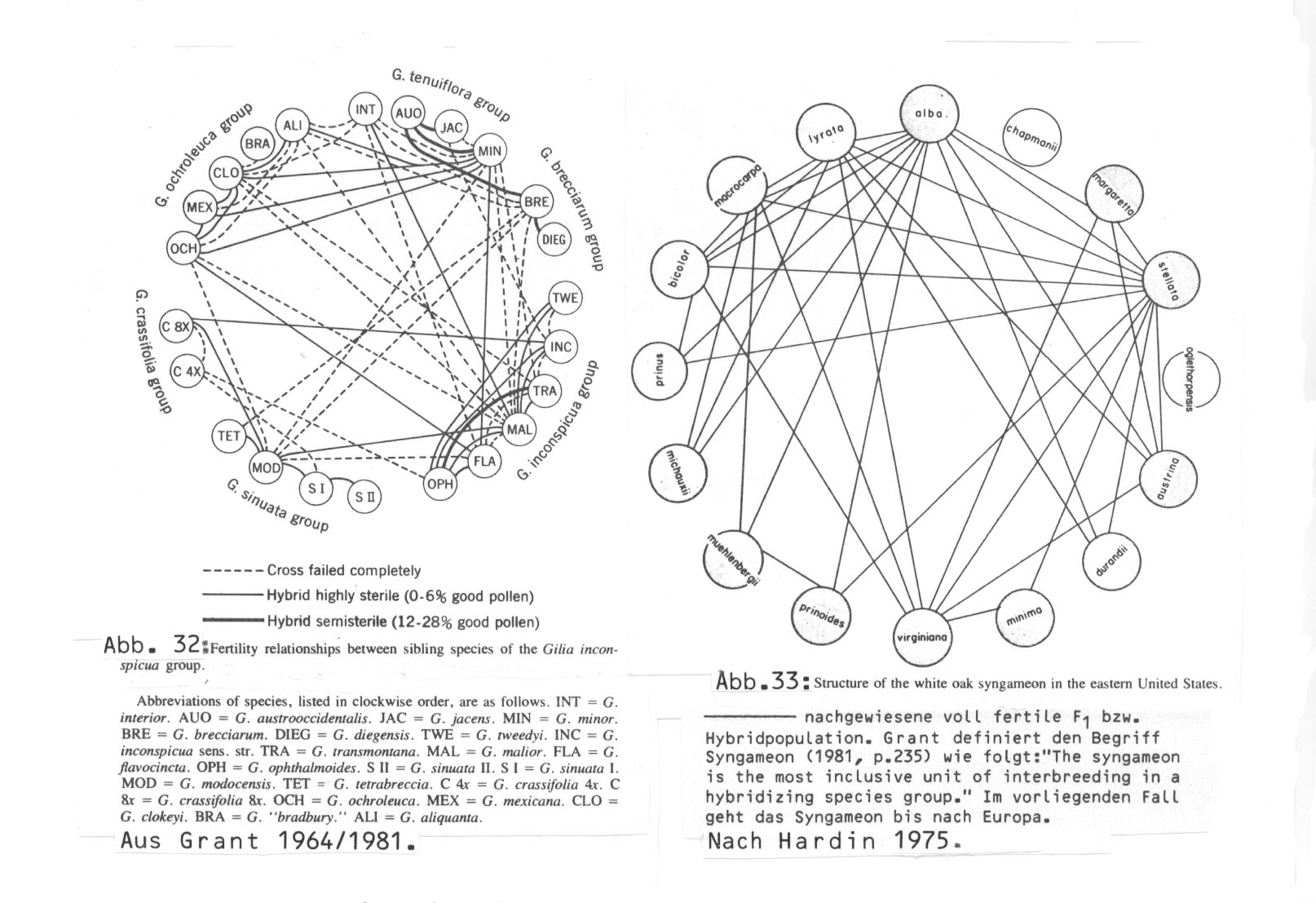
Zur Veranschaulichung diene der folgende Vergleich zwischen einem einjährigen Kraut (Gilia) und einem Baum (Quercus - 'white oak syngameon') (beide Abbildungen ebenfalls nach Grant pp. 144 und 240; verkleinert.)
Was bedeuten nun diese Ausführungen für unsere Frage nach dem Artbegriff?
Ich habe schon wiederholt darauf aufmerksam gemacht, dass die Synthetische Evolutionstheorie großes Interesse daran hat, Spezies in statu nascendi zu demonstrieren, um mit solcher Artbildung das Prinzip des Ursprungs aller Lebensformen auf unserer Erde zu implizieren. Zu welchen logischen und praktischen Schwierigkeiten die oben aufgeführten prä- und postzygotischen Mechanismen als Artbegrenzungskriterien führen können, habe ich anhand zahlreicher Beispiele gezeigt. Die zuletzt aufgeführten Beispiele der Kreuzungspolygone dürften diese Ausführungen weiter fundieren:
So kommen wir bei dem Geum-Beispiel aus den Schwierigkeiten mit der postzygotischen Isolation als Arttrennungskriterium gar nicht mehr heraus:
Die Hybriden aus Geum molle x G. aleppicum sind steril - also handelt es sich um zwei echte Arten. Die Nachkommen aus G. coccineum x G. molle sind voll fertil - also gehören die beiden systematischen Spezies nur zu einer Art. Da nun aber die Nachkommen aus G. aleppicum x G. coccineum partiell fertil sind, ist Genfluss sowohl von aleppicum zu molle als auch reziprok möglich (vgl. ebenso die weiteren 7 auf Seite 171 aufgelisteten Beispiele und ergänze die Liste). Die Frage erhebt sich, wie Sterilität sekundären (oft degenerativ-dysfunktionellen-pathologischen) Ursprungs im Artbegriff verarbeitet werden sollte.
Noch klarer stellt sich das Problem, wenn solche Sterilitätsbarrieren innerhalb von systematischen Arten auftreten, wie das zwischen Populationen der erwähnten Arten Layia glandulosa, Nigella arvensis und Elymus glaucus der Fall ist (an dieser Stelle sei auch wieder an das Reis- und Weizenbeispiel und die obigen Beispiele aus dem Tierreich erinnert).
Bei Nigella arvensis beispielsweise ist die F1 zwischen den Subspezies
(1) Nigella arvensis arvensis x N. arvensis glauca gut fertil, genauso bei
(2) N. arvensis glauca x N. arvensis brevifolia; die Kreuzung
(3) N. arvensis arvensis x N. arvensis brevifolia aber brachte
keinen Samenansatz. Nach einem Artbegriff, der undifferenziert postzygotische Isolationsmechanismen als Artabgrenzungskriterien gebraucht, hätten wir es im Falle von (1) und (2) mit nur einer Art zu tun, im Falle von (3) aber mit zwei verschiedenen Arten - obwohl über glauca voller Genfluss gewährleistet ist.
Bei Elymus glaucus müssten mehrere neue Arten aufgestellt werden, obwohl die morphologischen Unterschiede so gering sind, dass nicht einmal die Systematiker hier eine Serie verschiedener Arten unterschieden haben (vgl. die Abb. 34 auf der nächsten Seite).
Beim oben zitierten Mimulus-Beispiel müssten die voll fertilen interspezifischen Hybriden zur Zusammenfassung der Elternlinien zu einer Art führen (wie das auch bei vielen anderen Fällen bei den Kreuzungspolygonen durchgeführt werden müsste), während die intraspezifischen, aber vollig sterilen Nachkommen innerhalb von Mimulus laciniatus und M. nasutus zur Trennung der Elternlinien in verschiedene Arten führen müssten - die morphologischen Abgrenzungskriterien sind nicht kongruent mit den Fertilitätsverhältnissen. Der letztere Punkt zeigt sich auch bei allen drei von Grant aufgeführten Kategorien (vgl. Tab. 2, p. 175) und wird durch die Unterschiede der beiden ersten im Vergleich zur letzten noch unterstrichen. Zwar gehen die morphologischen Differenzen bei einjährigen Kräutern ungleich häufiger mit geringer Kreuzbarkeit einher als bei den übrigen Kategorien, dafür gehen bei ihnen aber auch umso häufiger Sterilitätsbarrieren mitten durch die Arten. Vergleichbare morphologische Differenzen gehen jedoch bei mehrjährigen Kräutern, Sträuchern und Bäumen häufig mit guter Kreuzbarkeit einher.
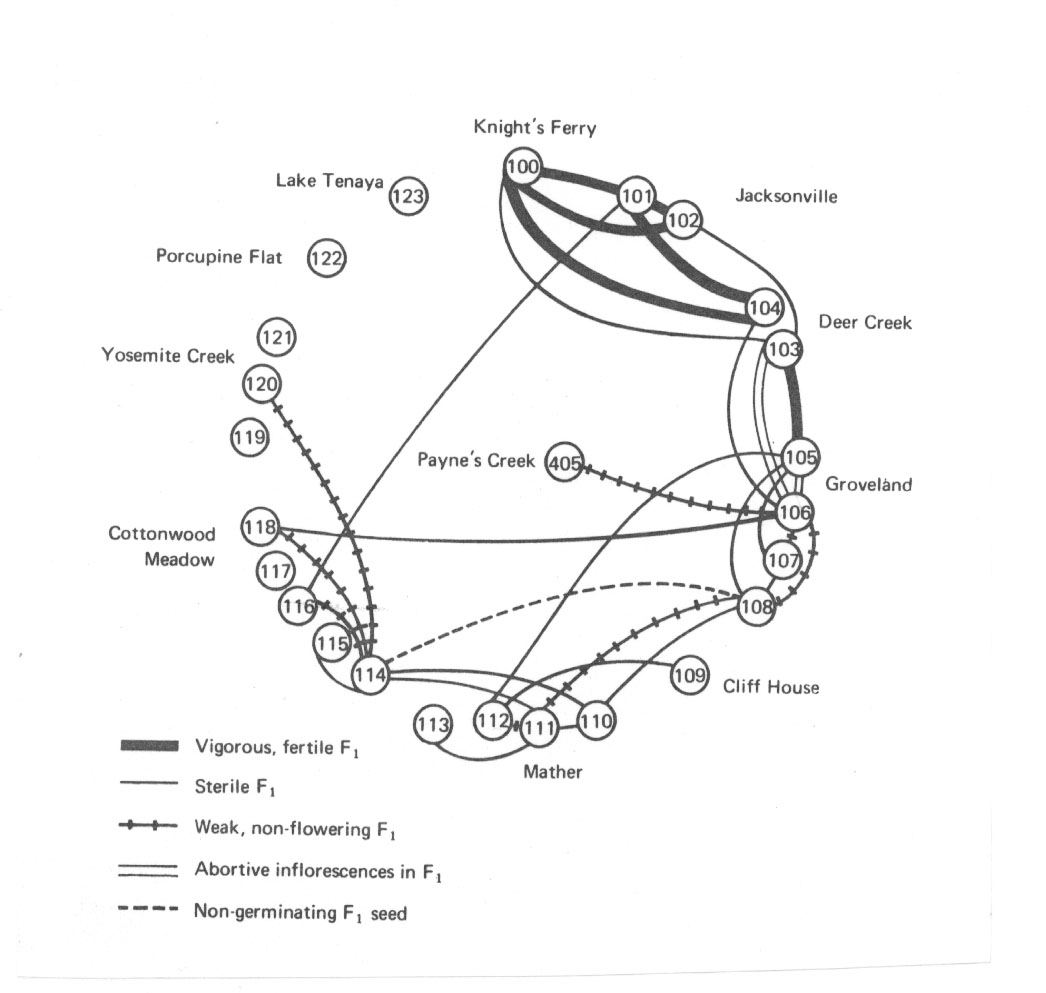
Abb. 34: Fertilitätsverhältnisse innerhalb der Elymus glaucus. Nach Snyder aus Briggs und Walters 1984.
Wie schon wiederholt gezeigt wurde, sind auch die Sterilitätsbarrieren in sich nicht einheitlich - weder auf genischer noch auf chromosomaler Ebene (auf dem ersterer Punkt beruhen ja die Schwierigkeiten bei Arten mit ziemlich einheitlicher Chromosomenstruktur). Auf chromosomaler Ebene können vergleichbare Strukturdifferenzen recht unterschiedliche Fertilitätsgrade zur Folge haben. Dazu einige konkrete Beispiele:
Hafner berichtet (1982) über die Kontaktzone zweier karyologisch definierter Taxa der zeltemachenden Fledermaus Uroderma bilobatum, wobei U. b. darisi 44 Chromosomen besitzt und U. b. convexum 38 Chromosomen (p. 861):
The chromosomal information (Baker 1981) indicates extensive hybrid production, including F1s and backcross individuals. Hence, introgression is documented on karyological criteria.
... Further, the distribution of karyotypes across the zone is examined, and does not depart significantly from that predicted by a model of simple diffusion of one parental form into the other. These results refute the statements that there is a paucity of F1s, a dearth of parental karyotypes, an overabundance of backcross individuals within the contact zone, and that speciation will ensue.
Die Unterschiedlichkeit im Karyotyp der beiden Linien hat also nicht zu der erwarteten Einschränkung in der Fertilität und zur 'Artbildung' geführt.
Carr schreibt (1980, p. 108) über die beiden Chromosomenrassen Tehama und Pauciflora der Asteracee Calycadenia pauciflora:
The results suggest that race Pauciflora is differentiated from Tehana by the equivalent of at least three chromosome translocations and the loss of one centromere, yet F1 hybrids have a mean fertility of over 40 per cent.
Die F1 Fertilität ist also trotz der starken Strukturheterozygotie bei Kreuzung dieser beiden aneuploiden Chromosomenrassen hoch.
Ladizinsky kommentiert die Tatsache, dass das Vicia sativa-Aggregat drei verschiedene Chromosomenzahlen enthält (nämlich 2n = 10, 2n = 12 und 2n = 14) und dass aus jeder Gruppe über karyotypische Variation berichtet worden ist (1981, p. 431):
Despite morphological and chromosomal differences the various entities are cross compatible and the F1 hybrids are partially fertile.
Außerdem sind in natürlichen Populationen Pflanzen mit unterschiedlichen Chromosomenzahlen Seite an Seite gefunden worden. F2 Populationen aus solchen Hybriden enthalten nach den bisherigen Untersuchungen zu etwa 15 % Individuen mit gleicher Chromosomenzahl, stabilem Karyotyp und guter Fertilität.
Wir erinnern uns auch an die Bemerkung von Herre und Röhrs (vgl. p. 33) von der 'bemerkenswerten Variabilität in den Chromosomenzahlen' bei den Haustieren, ohne dass dadurch eine Sterilitätsbarriere zustande käme.
Auf der anderen Seite können schon recht geringfügige Chromosomenunterschiede mit voller Sterilität einhergehen wie das oben aufgeführte Madia-Kreuzungspolygon samt der Liste ähnlich strukturierter Genera zeigen.
Mitzuberücksichtigen ist auch, dass die Toleranzgrenzen für genische und chromosomale Veränderungen bei den verschiedenen Organismengruppen offensichtlich unterschiedlich sind. Feine Unterschiede in der Chromosomenstruktur (ohne erkennbare pathologische Auswirkungen auf die betroffenen Individuen oder Populationen) sind beim Menschen beispielsweise nachweisbar (vgl. z.B. Passarge 1979; Nand et al. 1981; Nielsen et al. 1982; Kleinbrecht 1983), sie sind jedoch in keiner Weise mit der im Pflanzenreich häufig vorgefundenen Chromosomen-Variabilität innerhalb von systematischen Arten vergleichbar (man denke nur daran, was bei Pflanzen häufig an Deletionen, Inversionen, Duplikationen und Translokationen noch verkraftet wird, aber auch Themen wie Haploidie und Polyploidie geben über die höheren Toleranzgrenzen im Pflanzenreich Auskunft. Mit den Worten Walbots (1985, p. 168): "Considerable evidence suggests that normal development in plants is more tolerant of genetic imbalance than animals."
Dobzhansky et al. stellen (1977, p. 202) fest, dass 'Pflanzen eine wesentlich geringere Komplexität aufweisen als Tiere' und kommentieren:
... when developmental patterns are relatively simple, the possibility that elements from two different patterns can be combined to make a functional intermediate is much greater than when they are highly complex.
- was wohl mit dazu beiträgt, dass Pflanzen desöfteren eine höhere Toleranzgrenze bei Chromosomen- und Genom-Mutationen aufweisen als Tiere (vgl. die obigen Beispiele p. 158 und die Zitate aus Gottschalk 1976 und 1984).
Es gibt jedoch auch unterschiedliche Toleranzgrenzen innerhalb des Tier- und Pflanzenreichs. Sieht man sich neuere Arbeiten zum Thema Polyploidie im Tierreich näher an, so stellt man fest, dass Genom-Mutationen bei den Fischen und Amphibien ungleich häufiger verkraftet werden als bei den Reptilien (vgl. Ohno 1970; Lewis 1980; Therman 1980; Sessions 1982). Über die Säugetiere findet sich in der großen Arbeit von Lewis kein Kapitel, weil bislang keine Eupolyploidie in der Natur bei dieser Gruppe festgestellt werden konnte.
Lokki und Saura schätzen (1980, p. 277) die Zahl der Insektenarten auf 2,5 bis 3 Millionen, wovon etwa ein Drittel beschrieben seien und fahren fort:
In comparison with this diversity the list of known polyploid insect forms is exceedingly small, less than one hundred.
... In contrast to the apparent success of polyploid plants, polyploid insects represent rare exceptions, as the vast majority are diploid.
... All well documented polyploid insects reproduce partheno-genetically.
Zwei Fälle von Pseudogamie waren bis dahin bekannt. Im übrigen handelt es sich bei diesen polyploiden Insekten meist um flugunfähige Formen. Auch im Pflanzenreich sind starke Unterschiede in der Toleranz von Genommutationen konstatiert worden. "In summary ... polyploidy in gymnosperms is rare", stellt Delevoryas 1980, p. 216 nach Diskussion verschiedener Beispiele fest. Sequoia sempervirens (Taxodiaceae) ist z.B. die bislang einzige bekannte hexaploide Gymnosperme (2n = 66). Nach Walbot (1985) kommt das Phänomen der Polyploidie bei nur etwa 5 % der Gymnospermen vor, ist jedoch bei den Farnen allgemein verbreitet, und bei den Angiospermen belaufen sich die Schätzungen auf 45 % der dikotyledonen und 55 % der monokotyledonen Pflanzen (welches Phänomen nach Nilsson u.a. genetisch bedingt ist).
Was bedeuten nun all diese Befunde für den Artbegriff?
Zunächst sei hervorgehoben, dass mit diesen Tatsachen nirgends 'Höherentwicklung' (Integration und Koadaptation neuer morphologisch-physiologischer Strukturen etwa zu neuen Organen und Organsystemen) zu verzeichnen ist. Das wären doch überzeugende Beispiele, wenn über geographische Variation in einem weiten Verbreitungsgebiet z.B. neue Organe aufgebaut würden. Das Gegenteil ist hingegen nachweislich häufig der Fall: Durch Funktionsausfall von Genen und der damit verbundenen physiologisch-anatomischen Strukturen können Organismen in einem weiten Verbreitungsgebiet unterschiedlich weit degeneriert sein ("regressive Evolution" - Höhlentiere, Inselbewohner, flugunfähige Vögel und Insekten etc.). Durch Funktionsausfall von Genen kommt es auch in zahlreichen Fällen zu sekundären Sterilitätsbarrieren (Epistasie). Wenn auch durch anatomisch-physiologisch weitgehend neutrale Veränderungen (wie Chromosomenumbauten - Beispiele siehe oben) Sterilitätsbarrieren aufgebaut werden können, so wird doch das eigentliche Ziel des Neodarwinismus, den Ursprung der Komplexität aller Lebensformen mit solcher 'Artentstehung' zu erklären in keinem Falle erreicht. Selbst Stebbins schreibt nach Diskussion von Beispielen aus dem Pflanzenreich (1977, p. 223): "Semispecies of plants are not always, and perhaps not usually, species in statu nascendi."
Zweitens führt jeder Versuch, die Sterilitätsbarriere bei nahe verwandten Linien für die Artabgrenzung zu verabsolutieren, zu der Kette von Schwierigkeiten und Ungereimtheiten, die ich oben im einzelnen aufgeführt und besprochen habe (uferlose Speziesmacherei durch Produktion Hunderttausender Zwillingsarten, Artentrennung mitten durch Kultursorten von Weizen, Gerste, Erbsen etc., Artabgrenzung quer durch sonst recht einheitliche Populationen des blauen Polarfuchses, Zusammenfassung von blauen und weißen Populationen aber zu einer 'Art' und ähnlichen Beispielen bei Mäusen etc., logische und systematische Schwierigkeiten bei den Kreuzungspolygonen und viele weitere Probleme.)
Das bedeutet, dass mindestens alle durch genische Degeneration, Chromosomen- und Genommutationen bedingten Sterilitätsbarrieren unter den Artbegriff subsumiert werden sollten (zumal die beiden letzteren in ihren Möglichkeiten artbedingt prädestiniert sind).
Drittens ist festzuhalten, dass durch die reproduktive Isolation häufig nur eine ursprünglich in vielen Genen rekombinierende weitverbreitete Population in genisch und geographisch kleinere Sektionen aufgespalten wird. Reproduktive Isolation ist demnach in vielen Fällen ein von den (anfangs oft größeren) Möglichkeiten der Rekombination physiologischer und morphologischer Merkmale unabhängiges und sekundäres Ereignis, wie die folgenden Rekombinationsquadrate veranschaulichen:
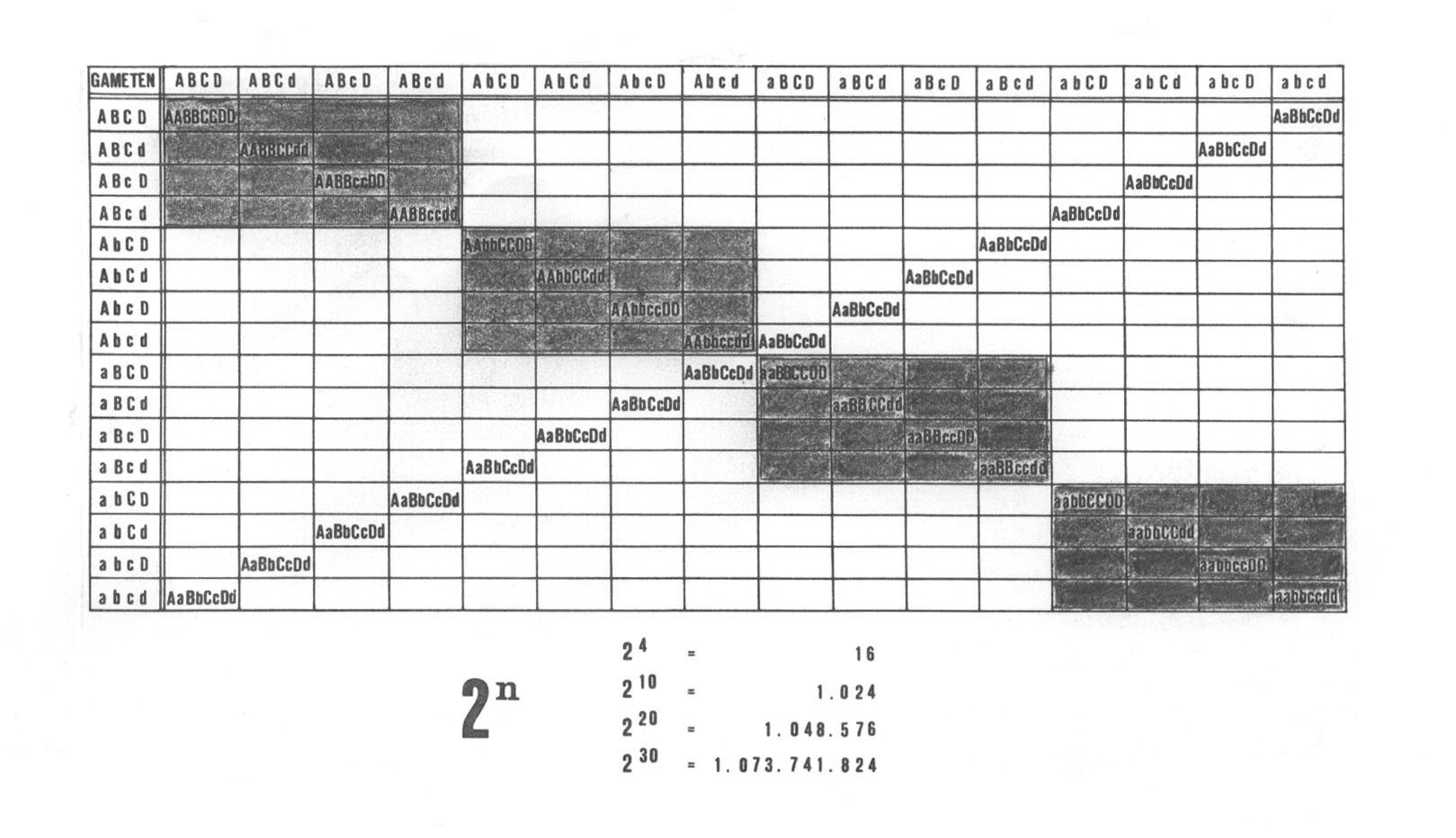
Abb. 35: Die ursprünglich 16 (stark unterschiedliche) morphologische Rekombinanten aufweisende Art P wird durch Dezimierung auf geographische Isolate unterschiedlicher ökologischer Struktur in 4 Subspezies mit geringerer Variation aufgespalten.
Die ursprünglich weit verbreitete in vielen Genen rekombinierende Art P zerfalle in die morphologischen Unterarten Pl, P2, P3 und P4 (der Einfachheit halber hier genetisch gleich groß dargestellt). Diese sollen kleinere Areale mit unterschiedlicher ökologischer Struktur besiedeln. Weitere für diese ökologischen Nischen nicht mehr unbedingt notwendige Genfunktionen können mit der Zeit durch Mutationen ausgeschaltet werden: Von den Phänotyp wesentlich mitbestimmenden Genen A bis D völlig unabhängig mutieren in den verschiedenen Populationen die Gene E - H, die sich nur oder fast nur physiologisch auswirken sollen. Die Subspezies sind optimal an ihren Lebensraum angepasst, aber durch Funktionsverluste nicht mehr so anpassungsfähig wie die Ausgangsart. Durch Mutationen der Gene E - H sowie Änderungen in der Chromosomenstruktur kann es zu folgenden Gruppen von Sterilitätsbarrieren kommen:
Reproduktive Isolation innerhalb der morphologischen Subspezies P1 - P4. Resultat: Zwillingsarten
Reproduktive Isolation zwischen den morphologischen Subspezies: Systematisch gute Arten mit Sterilitätsbarriere.
Reproduktive Isolation zwischen einer und den übrigen Subspezies (dann sind beispielsweise P1 - P3 weiterhin miteinander fertil, aber die F1 sei steril bei Kreuzungen mit P4.)
Kombination der Möglichkeiten 1 - 3: Reproduktive Isolation teilweise innerhalb einer morphologischen Subspezies und teilweise zwischen solchen. Als nächste Möglichkeit kann sich die Kreuzbarkeit mit fertilen Nachkommen auf Populationen zweier (oder mehrerer) morphologischer Subspezies erstrecken, welch letztere selbst jedoch reproduktive Isolation innerhalb ihrer Grenzen zu verzeichnen hat.
Schließlich könnte auch Nicht-Kreuzbarkeit (keine F1 möglich) mit einer (oder bei größeren Formenkreisen mehreren) Subspezies vorkommen.
In diesen Fällen sind wir davon ausgegangen, dass in den verschiedenen geographisch voneinander getrennten Subspezies die reproduktive Isolation verursachenden Mutationen unabhängig voneinander aufgetreten sind: In P1 beispielsweise ist E zu e und H zu h mutiert.Treten beide zugleich in einer F1 auf, so sei diese steril (Fall 1), ist e jedoch in P2 verbreitet und h in P3, dann haben wir reproduktive Isolation zwischen den Subspezies (Fall 2) etc.
Setzen wir weiter folgenden Fall:
In der Subspezies P1 sei (wie schon oben) E zu e mutiert, in P2 H zu h, in P3 G zu g.
Tritt Epistasie mit steriler F1 nur bei Kombination von e und h auf, beim Zusammentreffen von g und h jedoch nur in abgeschwächter Form, so dass partielle Fertilität in der F1 noch möglich ist und kommen keine negativen Interaktionen zwischen e und g vor, dann sehen die F1 en folgendermaßen aus:
|
(A) |
P1 x P3 |
fertil |
|
(B) |
P3 x P2 |
partiell fertil |
|
(C) |
P1 x P2 |
fertil |
(bei (C) vgl. die Geum-Beispiele p. 171)
Graphisch lässt sich die Situation folgendermaßen veranschaulichen:
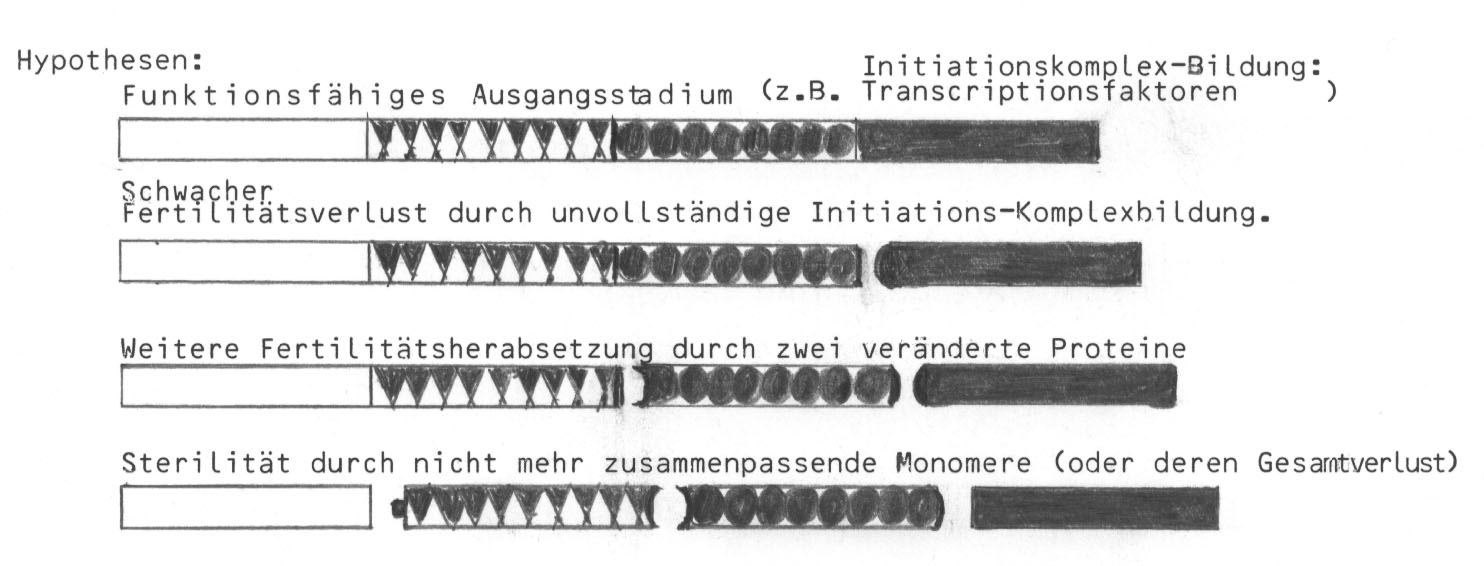
(Zum Thema Transcriptionsfaktoren vgl. z.B. S. F. Gilbert (1991): Developmental Biology, Sunderland, Mass; S. L. Wolfe (1993): Molecular and Cellular Biology, Belmont, Cal.) Auch an Strukturpolymere wäre zu denken. Vgl. p. 134.
Eine andere Möglichkeit wäre, dass Mutationen mit Fertilitäts-mindernder Wirkung schon in der ursprünglichen großen Population aufgetreten sind und nun mehr oder weniger zufällig über die inzwischen geographisch isolierten Subspezies verteilt sind. Der einfachen Veranschaulichung halber nehmen wir wieder unser Rekombinationsschema, diesmal mit den Genen J K L M, die zu j k L m mutiert seien. (Über die Homozygotenlinie des Rekombinationsquadrats hinaus gibt es selbstverständlich eine ganze Kette weiterer Möglichkeiten - einzeln oder in Kombination -, die aufzuführen noch mehrere Seiten beanspruchen würde.) Ich möchte an dieser Stelle nur das Prinzip veranschaulichen. Zu diesem Zwecke soll diese Rekombinantenserie auch mit der morphologischen Serie parallel laufen, obwohl das in der Realität sehr unwahrscheinlich ist.
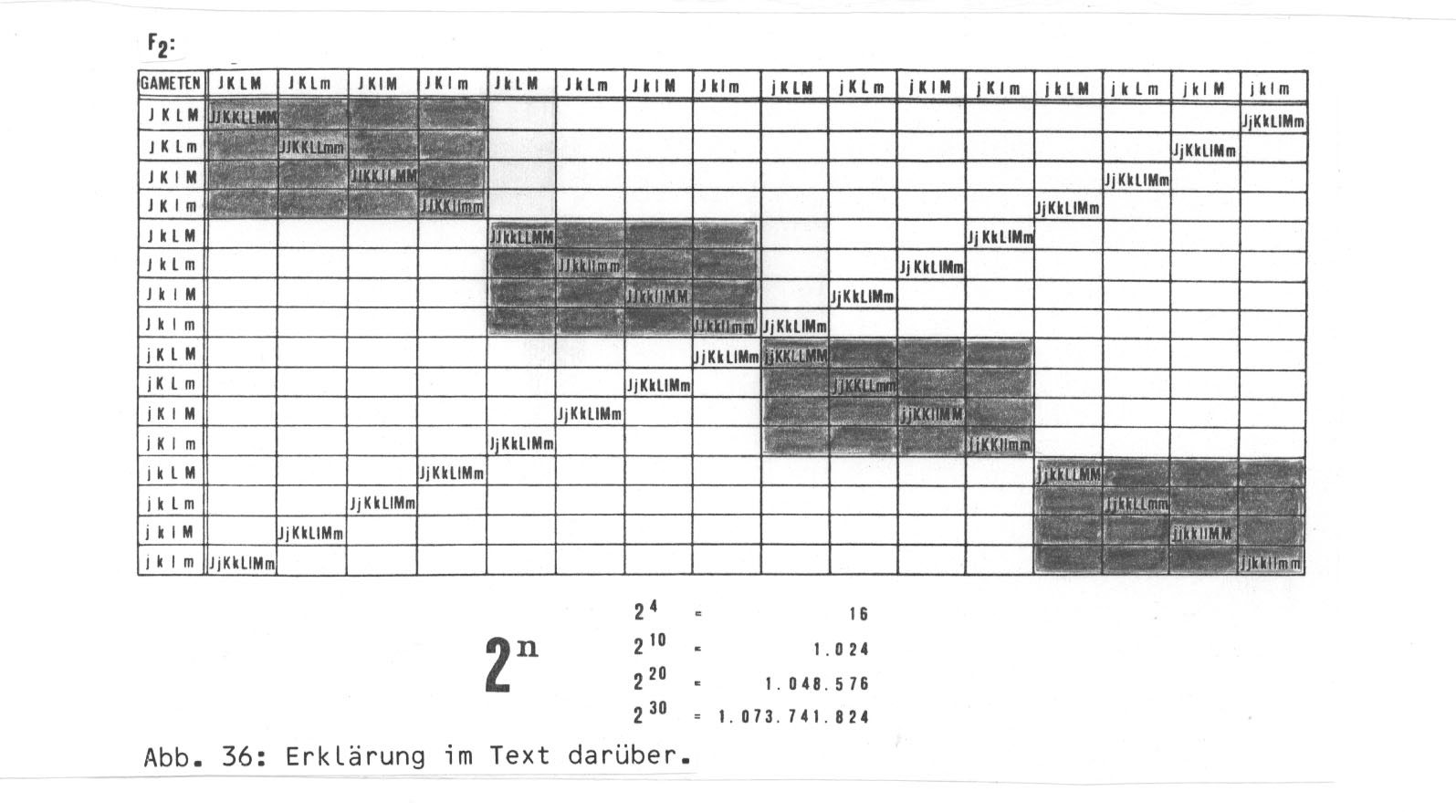
Abb. 36: Erklärung im Text darüber.
Bei Kombination der Allele m und n sei die F1 KK LL mm nn steril. Das kann z.B. eine Population aus dem morphologisch recht einheitlichen Genotyp AA BB CC DD betreffen (ideale Zwillingsarten).
Eine andere Möglichkeit wäre, dass beim Zusammentreffen von m und n keine Komplikationen auftreten, sondern erst bei Kombination der Gene ll mm nn bei den Subspezies P2 und P4. In diesem Falle hätten wir Zwillingsarten innerhalb von P2 und P4, dennoch würden zahlreiche Genotypen der anderen Subspezies mit Populationen von P2 und P4 fruchtbare Nachkommen erzeugen können (vgl. die Situation bei Mimulus zitiert p. 172). Im übrigen wäre die Rekombinante kk ll mm nn von P4 immer steril, was auch auf eine Reihe von Heterozygoten zutreffen würde.
Als weitere Möglichkeit soll erst beim Zusammentreffen von k, 1, m und n Epistasie auftreten. Dann gibt es eine Sterilitätsbarriere innerhalb von P4 (kk ll MM nn X kk ll mm NN --> F1 steril), aber auch mit Genotypen und Populationen von P1 bis P3 , und zwar jeweils die Hälfte der mit den Rekombinanten kk ll MM nn / kk ll mm NN gekreuzten Populationen. (In diesem Zusammenhang sei noch einmal auf die Mahnungen und Einschränkungen von Briggs und Walters p. 172 zum Thema Kreuzungspolygone hingewiesen.) Die Situation wird natürlich bei Berücksichtigung der Heterozygoten noch komplizierter.
Mehrere Autoren haben hervorgehoben, dass es "species-distinguishing genes" geben müsse (Lewontin 1974, p. 174 unter Hinweis auf Prakash), - das heißt, dass nur ein besonderer Teil des Genoms die Sterilitätsbarriere zwischen Zwillingsarten und anderen nah verwandten 'Arten' verursacht, während der größte Teil des Genoms unverändert bleibt. Der Autor schränkt (p. 164) allerdings ein, dass nur in ein paar Fällen "single gene substitutions with some modifiers" einen klaren 'Speziesunterschied' erklären und dass viel öfter mehrere Gen-Differenzen dafür notwendig seien, doch könne man derzeit unmöglich den genauen Teil des die Speziesdifferenz bedingenden Genoms bestimmen.
Einen Hinweis auf unsere obige Feststellung von sekundären Sterilitätsbarrieren findet sich beim selben Autor auf der Seite 179 seines Buches (wo verschiedene Beobachtungen zusammengefasst werden):
... where species are highly differentiated in their alleles there is at least a low-level polymorphism in one species for the genes that characterize the other. There is then a potential genetic transition between species that does not require the chance occurrence of new variation by mutations. That is, the overwhelming preponderance of genetic differences between closely related species is latent in the polymorphism existing within species.
In ähnlicher Weise schreibt Dobzhansky (1977, p. 194):
... given enough genetic divergence, superimposition of reproductive isolation may occur through a limited number of gene differences responsible particularly for isolating mechanisms. *
(Vgl. auch Mayr 1967, p. 299: "Morphologische Differenzierung scheint gewöhnlich weit schneller zu erfolgen als der Erwerb isolierender Mechanismen." - Was er auch für ökologische Differenzierung hervorhebt.)
Besonders hervorgehoben wird der folgende Punkt:
The genetic distances found between subspecies of the Drosophila willistoni group ... need not be the same as between subspecies in other groups of animals and plants. Let it be emphasized again that the processes of speciation occur in different ways in different organisms. The amount of genetic divergence that preceeds and follows the advent of reproductive isolation is not everywhere constant.
- worauf erwähnt wird, dass die genetische Distanz (in den Strukturgenen) zwischen Mensch und Schimpanse trotz der großen körperlichen und geistigen Unterschiede derjenigen bestimmter Zwillingsspezies von Drosophila-Arten entspricht, welch letztere "indistinguishable in external appearance" sind. Darauf beruht ja die vor allem mit King und Wilson (1975) einsetzende wissenschaftliche Kritik an der Idee, dass man morphologisch-anatomische und ethologische Differenzen mit der genetischen Distanz in Strukturgenen gleichsetzen kann (Nei glaubte 1972 sogar eine bestimmte, allgemein gültige Distanz zwischen Arten, Gattungen, Familien etc. berechnen zu können).
Schmidt fasst (1985, pp. 36/37) diese Frage wie folgt zusammen:
W.C. KING und A.C. WILSON (1975) untersuchten die genetischen Unterschiede zwischen Mensch und Schimpanse mit Hilfe von Proteinsequenzanalysen, immunologischen Methoden, Elektrophorese und Nukleinsäurehybridisation. Die Abweichungen waren so gering wie zwischen Geschwisterarten von Drosophila; die Übereinstimmung bei ihren vergleichenden Untersuchungen an 44 Proteinen betrug mehr als 99 %. Die gefundenen Abweichungen sind nach ihrer Ansicht viel zu klein, um die beträchtlichen Unterschiede in Anatomie, Physiologie und Verhalten von Mensch und Schimpanse befriedigend zu erklären. Demgegenüber waren z.B. die immunologischen Abweichungen der Albumine zweier Froscharten (Hyla und Rana) 20-30 mal größer als zwischen Mensch und Schimpanse, ebenso der mit elektrophoretischen Methoden festgestellte genetische Abstand zwischen verschiedenen Hyla-Arten, obwohl diese Froscharten sich in Anatomie und Physiologie weit näher stehen.
KING und WILSON vermuten deshalb, dass eine relativ kleine Zahl genetischer Veränderungen der Genregulation für die vergleichsweise großen Unterschiede in Anatomie und Verhalten zwischen Mensch und Schimpanse verantwortlich ist, von denen einige eher durch ein "Rearrangement" von Genen und Chromosomen verursacht sein dürften als durch Punktmutationen.
Auch dies darf man wohl mit aller Vorsicht als einen Hinweis darauf betrachten, daß uns noch eine ganze Dimension zum Verständnis der Vorgänge fehlt, die für die Evolution letztlich verantwortlich sind und daß es eine grobe Simplifizierung wäre, letztlich für alles nur Mutationen verantwortlich zu machen.
Allerdings unterscheidet sich hier die Situation von der der Geschwisterarten und Kreuzungspolygone insofern, als kein vernünftiger Biologe jemals Mensch und Schimpanse demselben 'Kreuzungspolygon' zuordnen würde. Hier geht es nicht um sekundäre Sterilitätsbarrieren, sondern um eine primäre Artabgrenzung auf (Regulator-)genetischer und plasmatischer sowie morphologisch-anatomischer und geistiger Basis. Dennoch zeigt dieses Beispiel besonders klar die mangelnde Korrelation zwischen physiologischen und morphologischen Daten, weshalb ich diesen Punkt an dieser Stelle mitaufgeführt habe.
Weitere Beispiele:
Carson schreibt über die morphologisch recht unterschiedlichen Drosophila-'Arten', die auf den neueren Vulkanen Hawais zu finden sind (1982, p. 18):
... with the exception of D. planitibia of Maui, considerable interspecific fertility exists among these species. This situation is especially striking in the partially sympatric pair of species silvestris and heteroneura. Furthermore, no hybrid sterility or postmating breakdown has been encountered in any interpopulation crosses within or between these two species, including crosses between Kona and Hilo types of D. silvestris (Craddoch, 1974; J.N. Ahearn, unpublished).
Die von Dobzhansky zitierten (morphologisch nicht unterscheidbaren) 'Zwillingsarten' sind durch eine Sterilitätsbarriere voneinander getrennt, die gut unterscheidbaren 'Arten' Hawaiis aber sind häufig miteinander voll fertil ("... more hybridizations have been achieved within this fauna than in any other group." - I.R. Bock 1984, p. 54). Durch eine Serie von Gen- und Chromosomenmutationen ließen sich nun zahlreiche Sterilitätsbarrieren innerhalb und zwischen den Drosophila-Arten Hawaiis aufbauen. Generell wird eine relativ junge Besiedlung Hawaiis durch viele Lebensformen veranschlagt, so dass meist die These vertreten wird, es sei noch nicht genug Zeit für eine häufigere Bildung von Sterilitätsbarrieren vergangen. Sicher spielt jedoch bei diesem Fragenkomplex die Zeit nicht die einzige Rolle.
In ihrer Arbeit THE CICHLID FISH OF CUATRO CIÉNEGAS, MEXICO: DIRECT EVIDENCE OF CONSPECIFICITY AMONG DISTINCT TROPHIC MORPHS (1982) beschreiben Kornfield et al. einen Fall von ökotrophisch und morphologisch stark differenzierten Formen, die jedoch zur selben Art gehören (p. 658):
By any phenetic criteria, the extreme distinctness of sympatric cichlids in Cuatro Ciénegas suggests the presence of two discrete species. A "small tooth" form with papilliform pharyngeal dentition feeds on plant material and possesses a relatively narrow head, slender pharyngeal jaw and long intestine. A "large tooth" form with molarform pharyngeal teeth feeds on snails and has a wider head, relatively stouter jaw and short intestine. The morphological and trophic differentiation between these two forms exceeds that observed among many closely related biological species and precisely mimics the differences which define numerous cichlids in the African Great Lakes.
Auch hier deutet sich wieder an, dass weitgehende morphologische Differenzen sekundären Sterilitätsbarrieren vorausgehen können. Die beiden von Kornfield et al. beschriebenen Formen gehören zu einer einzigen panmiktischen Population (und die Autoren weisen darauf hin, dass diese Formen ursprünglich auch als verschiedene Arten beschrieben worden sind und dass zahlreiche weitere Beispiele für solche ökologisch-morphologischen Divergenzen innerhalb panmiktischen Populationen existieren). Zu der Bemerkung der Autoren (p. 658): "If individuals from a panmictic population are recognized as species by standard taxonomic criteria, organismal diversity in nature may be more apparent than real." - ist hinzuzufügen, dass die organismische Diversität gewiß real ist, nur die daraus abgeleiteten Artenzahlen sind es nicht.
Auf der anderen Seite finden wir immer neue Beispiele für (struktur-) genetisch recht differenzierte Zwillingsarten. Adams et al. bemerken in ihrer Arbeit LARGE GENETIC DIFFERENCES BETWEEN SIBLING SPECIES OF BATS, Eptesicus, FROM AUSTRALIA (1982, p. 437):
What is clear is that even within mammals, morphological divergence and genetic divergence need not be correlated.
In den verschiedensten Beispielen zeigt sich immer wieder die Unabhängigkeit sekundärer Sterilitätsbarrieren vom übrigen morphologisch-anatomischen und oft auch physiologischen Divergenzgrad.
Kehren wir für ein weiteres Beispiel nach Hawaii zurück. Über die zur Kompositen-Familie gehörende Silberschwert-Gruppe berichten Carr und Kyhos (1981, p. 543):
The remarkable array of life forms within the silversword alliance exploits almost every conceivable terrestrial habitat in Hawaii... In spite of the spectacular and ecological diversity exhibited by the members of the silversword alliance, the occurrence of many natural intergeneric, intersubgeneric, and interspecific hybrids (Sherff, 1935, 1944; Kobayashi, 1973 b; and Carr 1978a) attests to the fact that they form a natural genetically cohesive group that has in all probability resulted from rapid evolutionary differentiation after a single colonization by a progenitor, possibly in less than 10 million years (...).
Was hier als "rapid evolutionary differentiation" bezeichnet wird, dürfte zum großen Teil auf Genmutationen, d.h. auf Ausfall bzw. Funktionsminderung von Genfunktionen zurückzuführen sein. Wird davon im Laufe der Zeit zunehmend auch noch jener Teil des Genoms betroffen, der Sterilitätsbarrieren innerhalb und außerhalb verschiedener Populationen verursachen kann, dann ließe sich im Sinne der obigen Rekombinationsquadrate eine Serie neuer durch solche Sterilitätsbarrieren voneinander getrennter 'Arten' benennen. Die Fragwürdigkeit eines solchen Unternehmens haben wir oben im Detail diskutiert. Wir wollen es nochmal hervorheben: Sekundäre und genisch wie chromosomal ungleichwertige (sowie teilweise reversible) Ereignisse mit den unterschiedlichsten Auswirkungen auf die Fertilität zweier Linien können nicht ohne die widersprüchlichsten Ergebnisse zum Maßstab für den Artbegriff gemacht werden. Sie müssen unter den Artbegriff subsumiert werden.
Zum Thema der Kreuzungspolygone seien abschließend die außerordentlich informationsreichen Studien von Vickery und Wullstein zitiert. Sie haben den Verwandtschaftsgrad der sechs systematischen Arten und einigen Varietäten der Section Erythranthe der Gattung Mimulus mit den unterschiedlichsten taxonomischen Ansätzen und Methoden erarbeitet. Wir folgen hier der Arbeit von Vickery (1984). Zuerst seien die Arten und Varietäten genannt (Nummern zur Identifizierung der Populationen in den Abbildungen):
Mimulus cardinalis Douglas
6651C Santa Catalina Mtns. Pina Co., Arizona, USA
7113C Los Trancos Creek, San Mateo Co., California, USA
7120C San Antonio Peak, Los Angeles Co., California
M. eastwoodiae Rydberg
6079E Bluff, San Juan Co., Utah, USA
M. lewisii Pursh.
5875L Albion Basin, Salt Lake Co., Utah
6103L Ice Lake, Placer Co., California
M. nelsonii Grant
6271N Devil's Backbone, Durango, Mexico
M. rupestris Greene
9102R Sierra de Tepotzlan, Morelos, Mexico
M. verbenaceus Greene
5924V Grand Canyon, Coconico Co., Arizona
Die Gruppe ist von Alaska bis nach Zentralmexiko und vom Pazifik bis zu den Rocky Mountains verbreitet. Die beiden Varietäten von Mimulus lewisii Pursh sind rosafarben, während alle anderen Spezies leuchtend rot blühen. Der Autor berichtet über seine und Wullsteins Studien (p. 9):
We studied the same set of nine populations using (1) numerical taxonomic, (2) chemotaxonomic, (3) comparative allozyme, (4) DNA/DNA hybridization and (5) standard cytogenetic approaches. The numerical analysis distinguished four groups... The chemotaxonomic investigation based on floral pigments delimited two groups. The comparative allozyme study also segregated two groups, but different ones. The DNA/DNA hybridization experiments based on fast annealing, repetitive DNA's grouped most of the section together except for three of the red-flowered taxa. Lastly, the cytogenetic study revealed n = 8 in all forms but a spectrum of F1 hybrid fertilities that revealed an interfertile group of five taxa. Overall, the five approaches yield results that are only partially congruent (Fig. 37). The two pink-flowered M. lewisii varieties form a distinctive group. The rare endemic, M. rupestris, is only loosely tied in to the complex.
Die nachstehend wiedergegebenen Abbildungen verdeutlichen die Ergebnisse im Detail (Abb. 37; aus Vickery 1984; Erklärung im Text):
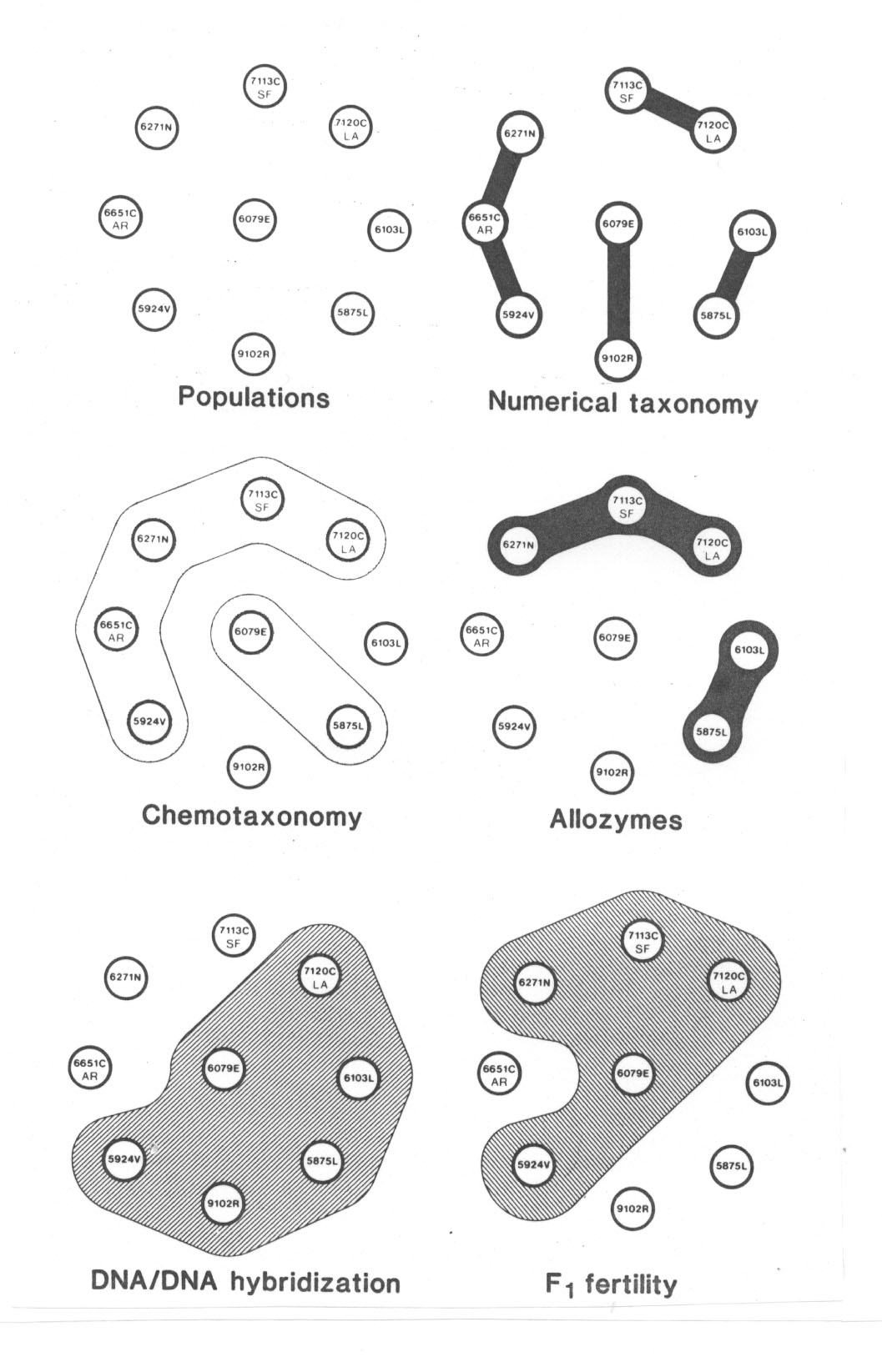
Wie zitiert, sagt Vickery zunächst sehr vorsichtig, dass die fünf Methoden nur partiell kongruente Resultate liefern. Sieht man sich die Ergebnisse genauer an, so stellt man fest, dass keine der fünf Methoden mit den morphologischen Artabgrenzungen ganz übereinstimmt. So ist z.B. die Population 6651 C von Mimulus cardinalis in vier der fünf Ansätze von den beiden anderen Populationen derselben Art getrennt (einschließlich F1-Fertilität), nur chemotaxonomisch gehört sie zur selben Art.
Nach den verschiedenen Untersuchungsmethoden könnte man folgende Artabgrenzungen vornehmen:
Numerische Taxonomie:
|
Spezies A umfasst: |
Mimulus cardinalis, die Populationen 7113 C und 7120 C |
|
|
|
|
Spezies B: |
M. lewisii, beide Populationen |
|
|
|
|
Spezies C: |
M. eastwoodiae |
|
|
M. rupestris |
|
|
|
|
Spezies D: |
M. cardinalis, Population 6651 C |
|
|
M. nelsonii |
|
|
M. verbenaceus |
Chemotaxonomie:
|
Spezies A umfasst: |
M. cardinalis, alle drei Populationen |
|
|
M. nelsonii |
|
|
M. verbenaceus |
|
|
|
|
Spezies B: |
M. eastwoodiae |
|
|
M. lewisii, Population 5875 L |
|
|
|
|
Spezies C: |
M. lewisii, Population 6103 L |
|
|
|
|
Spezies D: |
M. rupestris |
Allozyme:
|
Spezies A umfasst: |
M. cardinalis, Populationen 7113 C und 7120 C |
|
|
M. nelsonii |
|
|
|
|
Spezies B: |
M. lewisii, beide Populationen |
|
|
|
|
Spezies C: |
M. cardinalis, Population 6651 C |
|
|
|
|
Spezies D: |
M. verbenaceus |
|
|
|
|
Spezies E: |
M. rupestris |
|
|
|
|
Spezies F: |
M. eastwoodiae |
DNA/DNA Hybridisation:
|
Spezies A umfasst: |
M. cardinalis, Population 7120 C |
|
|
M. lewisii, beide Populationen |
|
|
M. rupestris |
|
|
M. verbenaceus |
|
|
M. eastwoodiae |
|
|
|
|
Spezies B: |
M. cardinalis, Population 6651 C |
|
|
|
|
Spezies C: |
M. cardinalis, Population 7113 C |
|
|
|
|
Spezies D: |
M. nelsonii |
F1-Fertilität:
|
Spezies A umfasst: |
M. nelsonii |
|
|
M. cardinalis, Populationen 7113 C und 7120 C |
|
|
M. eastwoodiae |
|
|
M. verbenaceus |
|
|
|
|
Spezies B: |
M. cardinalis, Population 6651 C |
|
|
|
|
Spezies C: |
M. lewisii, Population 6103 L |
|
|
|
|
Spezies D: |
M. lewisii, Population 5875 L |
|
|
|
|
Spezies E: |
M. rupestris |
Nun bedarf es keiner langen Diskussion, dass die Übersichtlichkeit in der Systematik vollends verloren ginge, wenn man jede der Methoden verabsolutieren und entsprechende Artabgrenzungen und Benennungen vornehmen würde. Es gäbe dann jeweils sechs verschiedene Schlüssel zur Artbestimmung, die je nach Interesse und Neigung des einzelnen Forschers benutzt würden (und weitere Methoden wie die der DNA-Sequenzierung dürften noch hinzukommen).
Vickery ist nun der Auffassung, dass sich die verschiedenen Ergebnisse gar nicht widersprechen, sondern einander ergänzen, wenn er (p. 11) schreibt:
In a sense each approach views the complex from its own perspective. The numerical approach "sees" phenetic similarity, possibly the result of a common evolutionary history and of similar environmental pressures. The chemotaxonomic approach "sees" the floral pigment differences and probably reflects differing pollination systems as well. The comparative allozymes may reflect common descent and similar environmental pressures also. The DNA/DNA hybridization results "see" the similarity of fast-annealing DNA, i.e. presumably of regulatory genes and suggest close, basic genetic similarity. The cytogenetic results "see" the location of barriers to gene exchange. Each approach adds to our information about the biology of the group - often permitting distinctions to be drawn between entities that other approaches did not resolve. Clearly, they all are complementary. Therefore, it seems fair to conclude that they all can be thought of as part of biosystematics in the broad sense.
Wir stimmen mit dieser Deutung grundsätzlich überein. Vickery hat die Komplementaritäts-Interpretation in folgender Abbildung zusammengefasst (p. 13):
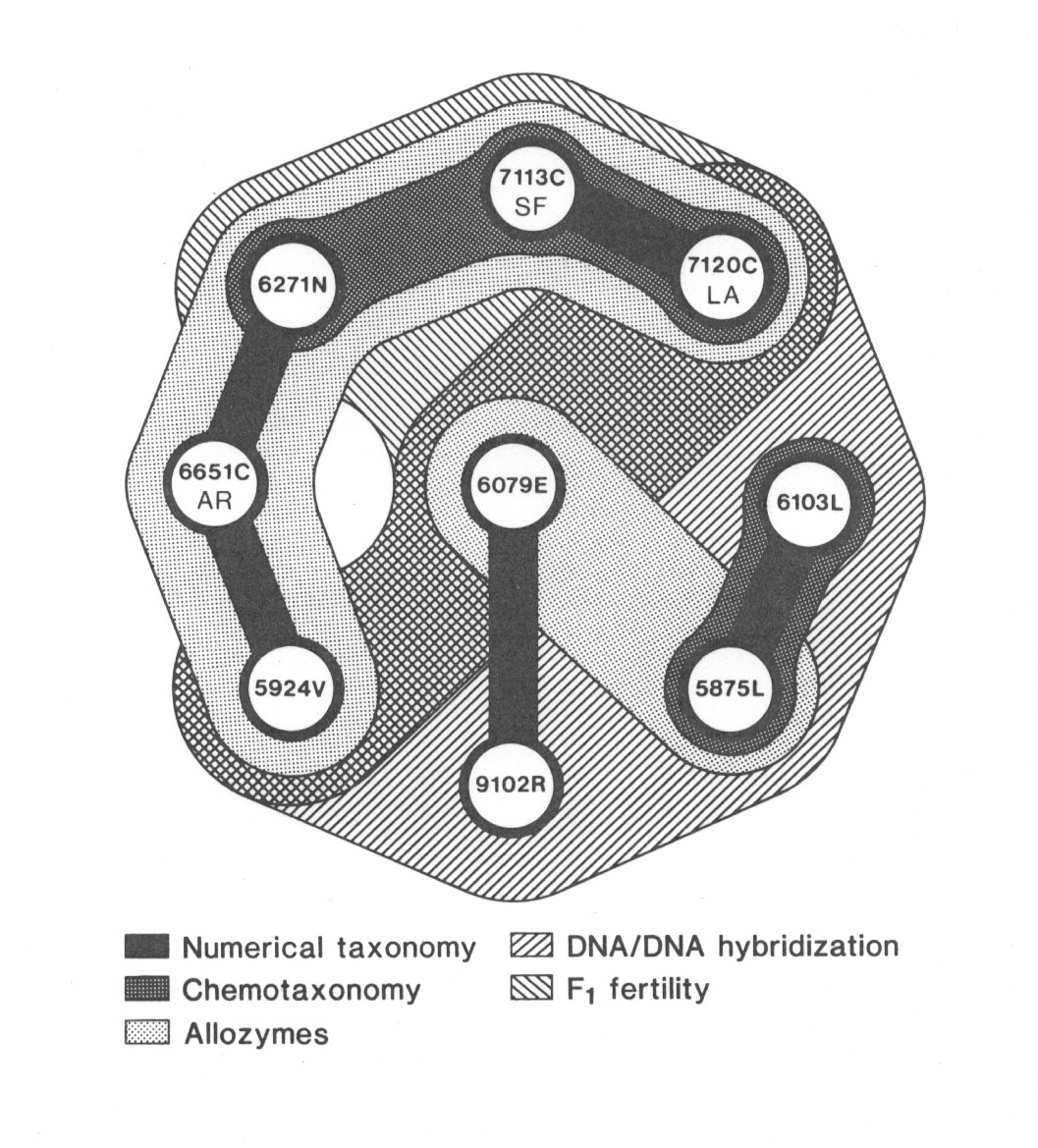
Abb. 38: Comparative groupings of the nine taxa of section Erythranthe by the 5 different approaches to classification. Aus Vickery 1984.
Die Frage nach dem Artbegriff und der Richtigkeit der Artabgrenzungen stellt der Autor in diesem Zusammenhang nicht. Jede Artabgrenzung innerhalb der (Mimulus-)Formen wäre auch insofern Willkür als eine Entscheidung zugunsten einer Methode alle anderen an verschiedenen Stellen grundlos ignorieren müsste. Im Sinne des Komplementaritäts-Verständnisses gibt es nur eine Lösung, die verschiedenen Ansätze auf einen gemeinsamen Nenner zu bringen, nämlich die Zusammenfassung der 6 morphologischen Arten (einschließlich ihrer Varietäten) zu einer Art. Innerhalb dieser Art lassen sich dann - je nach taxonomischer Methode - unterschiedliche Gruppierungen vornehmen: morphologische, numerisch-taxonomische, chemotaxonomische, Allozym-, DNA/DNA Hybridisations- und F1-Fertilitäts-Gruppen.
Weiter lässt sich im Anschluss an unsere Rekombinationsquadrate und im Sinne von Ohnos 'Dispensable Gene'-Konzept (1985, p. 164: "... the notion that all the still functioning genes in the genome ought to be indispensable for the wellbeing of the host should be abandoned once and for all.") sowie unter Berücksichtigung der Mutationsrate und genetischen Bürde folgende Deutung für die Bildung der unterschiedlichen Gruppen geben:
Die (alle Gruppen) umfassende Art verfügte ursprünglich über ein ziemlich gleichartiges Genpotential von Alaska bis nach Mexiko und hat mit der Zeit und unter den verschiedensten Umständen einen Abbau (und Umbau) des anfangs reichhaltigeren genetischen Potentials mit Auswirkungen auf den unterschiedlichen Ebenen erfahren.
Besonders deutlich erscheint die Situation bei der rosablühenden Mimulus lewisii. Die Blütenfarbe weist bereits auf den Abbau (bzw. Blockierung) genetischer Strukturen hin. Die auf der Funktion vieler Gene beruhende Anthozyansynthese kann durch Ausfall (Punktmutation/Deletion) oder Blockierung (Transposons) zu einer ganzen Palette unterschiedlicher Blütenfarben führen (die in Revertanten durch Veränderung der DNA-Sequenz noch weiter ausgebaut sein kann - vgl. p. 457).
Lamprecht hat das (1974, pp. 126-143) für Pisum wie folgt erarbeitet (es sei noch einmal hervorgehoben, dass alle kleinen Buchstaben für biochemisch meist durch Funktionsausfall gekennzeichnete rezessive Allele stehen - vgl. Zitate von Watson und Kaudewitz p. 123.):
|
Purpureus, Purpurfarbig (Rotblütig), Dull Dusky Purple |
: A Am Ar B Cr Ce |
|
Flavopurpureus |
: A Am ? B ? ? Paf |
|
Coeruleoviolaceus, Blauviolett, Nigrosinviolett |
: A Am ar B Cr Ce |
|
Clariroseus, Rosa, Lathyrusrosa, Deep Rose Pink |
: A Am Ar b Cr Ce |
|
Roseialbus, Blassrosa, Pale Rose Pink |
: A Am ar b Cr Ce |
|
Albus, Weiß, White |
a (mit beliebigen Kombinationen der übrigen Gene für Blütenfarbe); am: gleiche Wirkung wie a, aber bisweilen mit ganz blass rosa Flügel. |
|
Fuscopurpureus, Bräunlichpurpur, Indian Lake |
: A Am Ar B cr Ce |
|
Roseus, Rosa, Rose Color, Cerise |
: A Am Ar B Cr ce |
|
Palleopurpurascens, Blass Rosapurpur, Pale Rose-Purple |
: A Am Ar B cr ce oder : A am Ar B Cr Ce |
|
Malvaceus, Hell Malven, Light Mauve |
: A Am ar B cr Ce |
|
Antiqueroseus, Antik Rosa, Old Rose |
: A Am Ar b cr Ce |
|
Begoniaerubrum, Begonienrot, Rouge pivoine |
: A Am Ar b Cr Ce beg |
|
Rubicundus, Matt Rosenrot, Dunkelmatt Geraniumrot, Rose Red |
: A Am Ar b Cr Ce rub |
|
Salmochraceus, Hell Matt Ockergelb mit zartem Anflug von etwas Lachsfarben, Gelblich Fleischfarbig (Bei Pisum arvense oect. fulvum festgestellt) |
: A Am ar b cr ce Cit Cm |
"Die Kombination der beiden Gene Cit und Cm mit Ar, B, Cr und Ce (s.o.) gibt 64 verscheidene genotypische Konstitutionen. Hinzu kommt noch, dass, wie die Blütenfarbe der F1 zeigt, Heterozygotie in den Genen Cit und Cm, oder wenigstens in einem dieser beiden, intermediäre Farben bedingt. Salmochraceus wurde oben schon beschrieben. Unten folgt die Beschreibung von elf Blütenfarben, die konstant erhalten worden sind. Diese bilden natürlich nur einen Teil der ausgespaltenen neuen Blütenfarben. Sie geben aber ein gutes Bild von dem Reichtum an verschiedenen Farben und Nuancen, die durch die genannte Kombination erhalten werden können" (Lamprecht 1974, p. 137), - worauf die Blütenfarben Clariluteus, Clarimalvaceus, Claricorallinus, Corallinus, Pallidosanguineus, Sanguineus, Nubilorufus, Ochraceorubidus, Rubroviolaceus, Nubiloroseus und Nubilcorallinus detailliert beschrieben werden.
Man kann an diesem Beispiel besonders deutlich das Prinzip der Degeneration genetischer Strukturen mit der Begleiterscheinung erhöhter Variabilität erkennen: die Pisum-Ausgangspopulation besaß das gesamte dominante Genpotential, und durch Mutation und Isolation gingen im Laufe der Zeit bei verschiedenen Populationen (sowohl in der Natur als auch verstärkt unter Kulturbedingungen) die Funktion verschiedener (jetzt rezessiver) Gene verloren.
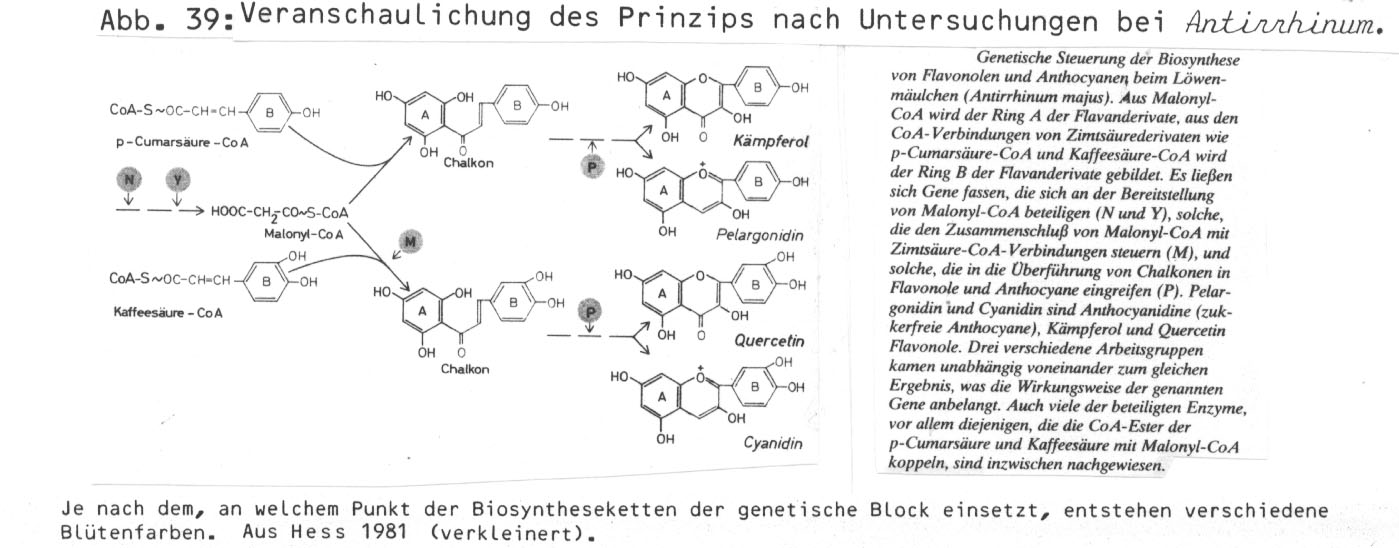
(Details zum Anthozyansystem vgl. z.B. Harte 1974, Hahlbrock 1981, Kochs und Griesebach 1986; bei den Autoren weitere Literatur.)
Das gleiche trifft im Prinzip auch auf die Synthesen vieler anderer Stoffklassen zu. Übrigens demonstrieren uns beispielsweise die Laubblätter in den gemäßigten Zonen jeden Herbst aufs neue, wie durch Strukturabbau Variabilität erzeugt werden kann. Durch den Abbau des Chlorophylls werden aber offenbar auch weitere biochemische Reaktionen in Gang gebracht. "The bright colors of leaves come about when large quantities of colorless flavonols are converted into anthocyanins as chlorophyll breaks down" (Raven und Evert 1981, p. 393). Störungen des normalen Metabolismus können die Anthozyanbildung fördern: "Zuckeranreicherung bewirkt oft eine merkliche Förderung der Anthocyanbildung: geringelte Zweige des Rebe oder des Hartriegels ergeben deshalb infolge der Unterbrechung der Zuckerableitung alsbald eine blutrote Herbstfärbung" (von Denffer 1979, p. 69).
Darüber hinaus wird mit der Entdeckung immer neuer Pseudogene, 'fossiler' und 'stiller' Gene die Blockierung bzw. der Abbau genetischer Strukturen auf molekulargenetischer Ebene zunehmend klarer (auch wenn ein Teil der Pseudogene auf Genduplikationen zurückgeht). Wegen der Replikationsgenauigkeit durch die verschiedenen pro- und eukaryontischen Poplymerasen und Reparatursysteme wird von Ohno (1985) die Halbwertzeit funktionsloser Gene auf Millionen von Jahren datiert (bei Fischen etwa 50 Mill. Jahre). Auf das Thema Pseudogene etc. wollen wir im Mutationskapitel zurückkommen.