NACHTRÄGE 1999 und 2002
NACHTRAG 1999
In den letzten zehn Jahren ist sehr viel auf dem Gebiet der Evolutionsforschung geschehen. Zu unserer Thematik, der Entstehung des Auges, beispielsweise, zeichnet sich die bisher größte "Revolution" seit rund 100 Jahren unter evolutionstheoretisch orientierten Biologen ab: Als Tatsache (und damit unumstößlich und vollkommen sicher) galt praktisch allen Theoretikern, dass "das Auge" vielfach unabhängig voneinander durch Mutation und Selektion entstanden sei (siehe Einleitung), jetzt allerdings glaubt eine nicht unbeachtliche Gruppe von Forschern, Beweise dafür gefunden zu haben, dass sämtliche der postulierten Entwicklungslinien des Auges doch auf einen gemeinsamen Vorfahren zurückzuführen seien. (Science 265:785-789,1994; Science 267: 1788-1792, 1995, sowie Beitrag zu Duboules Guidebook to the Homeobox Genes 1994).
Als wichtigste 'Message' der neueren Auffassungen für die Evolutionsfrage erscheinen mir vor allem die folgenden Aussagen:
"Because Pax-6 is involved in the genetic control of eye morphogenesis in both mammals and insects, the traditional view (10) that the vertebrate eye and the compound eye of insects evolved independently has to be reconsidered" (Quiring et al. 1994, p.785). Or p. 788:"...we have detected DNA sequences similar to those of Pax-6 in flatworms (Dugesia tigrina) (30) and in nemerteans (31), which are among the most primitive metazoa with eyes (28). If the corresponding genes in flatworms and nemerteans are also involved in eye morphogenesis, the concept that the eyes of invertebrates have evolved completely independently from the vertebrate eye has to be reexamined. Also, the hypothesis that the eye of cephalopods has evolved by convergence with the vertebrate eye (10) is challenged by our recent finding of Pax-6 related sequences in the squid Loligo vulgaris. (32)"
Das steht nun im klaren Kontrast zu Salvini-Plawen und Mayr (als vielfach zitiertes Musterbeispiel für die bis vor kurzem vorherrschende Überzeugung von der vielfachen Konvergenz der Entstehung von Lichtsinnesorganen):
"As we shall show, photoreceptors of various degrees of differentiation have been evolved independently in at least 40, if not 65 or even more separate phyletic lines" (1977, p. 209; - Hervorhebungen im Schriftbild von mir).
Siehe die ausführliche Begründung der Verfasser in ihrem Beitrag. Demnach sollten bei den verschiedenen Tiergruppen allein die Linsenaugen mindestens 15mal unabhängig voneinander entstanden sein (Salvini-Plawen und Mayr, S. 255).
Im Rahmen der Evolutionstheorie müssten hingegen nach den neuen Ergebnissen und Interpretationen Wirbeltiere, Insekten, Ascidien, Cepalopoden, Nemertinen und andere Tiergruppen einen gemeinsamen Vorfahren gehabt haben, der bereits die (gemeinsamen) homöotischen Hauptregulatorgene und auch dazugehörende (für die Bildung von vielleicht noch einfachen Lichtsinnesorganen) Targetgene aufzuweisen hatte.
Hier erhebt sich nun die Frage: Wer oder was soll dieser gemeinsame Vorfahr oder diese gemeinsame Ausgangsform gewesen sein?
Zur Herkunft der Chordaten schreibt W. Gehring zusammen mit R. Wehner (Lehrbuch der Zoologie 1990, p. 710):
"Der evolutive Ursprung der Chordaten liegt völlig im Dunkeln." Und über die Tunicata (Ascidien, p. 747): "Die am Vorderende des laryngealen Neuralrohrs liegenden Sinnesorgane können dagegen nicht mit den Augen und Labyrinthorganen der Wirbeltiere homologisiert werden." Zu den Acraniern (p. 749): "Auch für die längs des Rückenmarks angeordneten Pigmentbecherocellen gibt es bei den Vertebraten keine Entsprechung. Ebenfalls im Gegensatz zu Wirbeltieren steht das Fehlen von Augen und Schweresinnesorganen." Erst die Agnatha weisen typische Wirbeltieraugen auf: "Die bei den Larven (der Neunaugen) unter der Haut liegenden Blasenaugen entwickeln sich zu typischen Vertebratenaugen " (p. 752) (Hervorhebungen im Schriftbild von mir).
Nach den neuesten Arbeiten kann ich wohl davon ausgehen, dass hier in künftigen Auflagen vermutlich einige Korrekturen vorgenommen werden.
Zur derzeit noch weit verbreiteten Pterobranchier-Hypothese des Ursprungs der Chordaten führen Wehner und Gehring einige Einwände auf (S. 710), erwähnen dann die "Wurm"- oder Hydroskelett-Hypothese und zeigen dazu ebenfalls einige Schwierigkeiten (beide Segmentierungssysteme wohl unabhängig(?) voneinander entstanden etc.). Zur "Wurm-Hypothese" haben Romer und Parsons recht ausführlich die Hauptschwierigkeiten zusammengefasst, und ich möchte den daran interessierten Leser auf die Ausführungen der Verfasser verweisen (1983, S. 34-36, Vergleichende Anatomie der Wirbeltiere).
Bleibt man innerhalb phylogenetischer Auffassungen, so erscheint die von Romer und Parsons vertretene Pterobranchier-Hypothese wahrscheinlicher. "Of all the possibilities, most now converge on seeing the pterobranch hemichordates as closest to the origins of the deuterostomes..." Willmer 1990, p. 325. Die Pterobranchier aber haben keine Augen!
Salvini-Plawen und Mayr führen folgende Lebensformen auf, die nach Auffassung der Autoren wohl meist primär keine Augen besitzen: Anthozoa (Cnidaria), Cestoda (Plathelminthes), Gnathostomulida, Acanthocephala (Nemathelmintes), Niedere Mollusken (Caudofoveata, Solenogastres, Tryblidiida, Scaphopoda), Priapulida, Myzostomida, Phoronidea (Lophophorata), Pterobranchia (Branchiotremata), drei der fünf Gruppen der Echinodermata (Crinoidea, Echinoidea, Ophiuroidea), Larvacea (= Copelata; Tunicata).
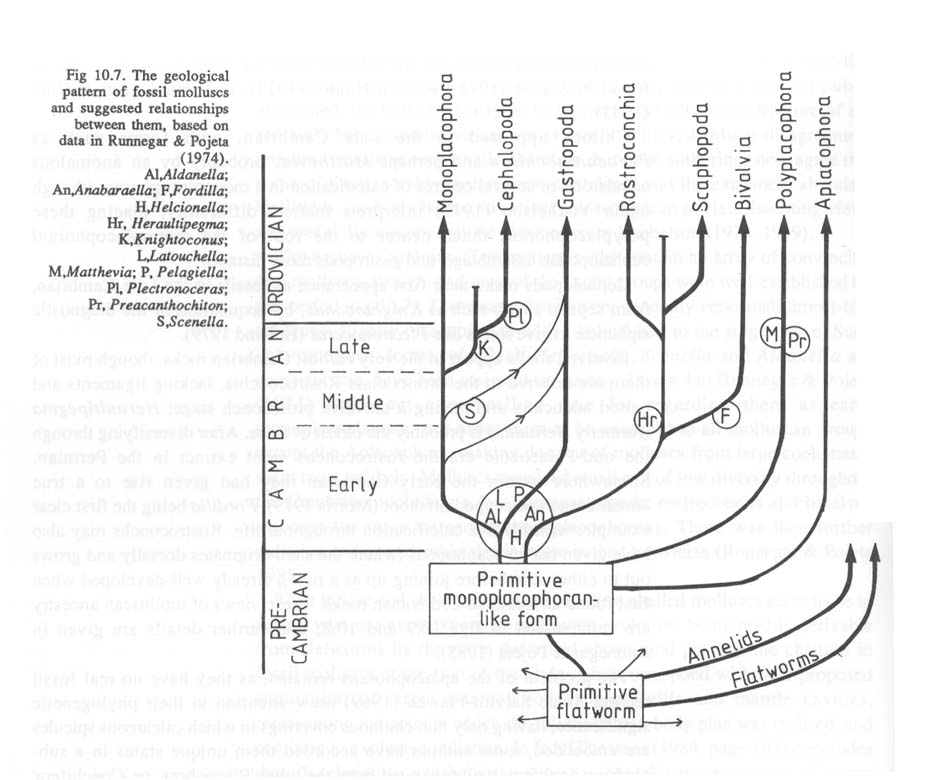
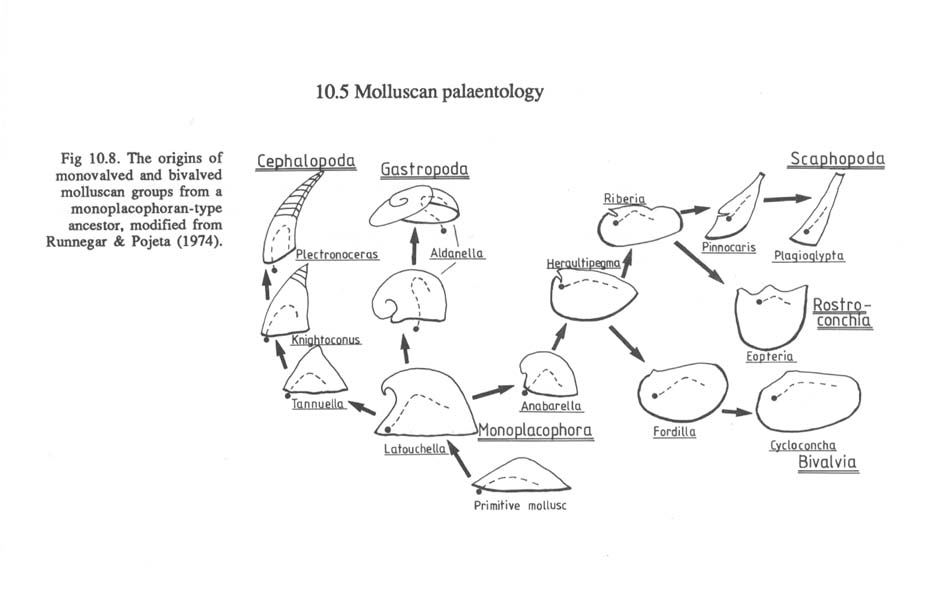
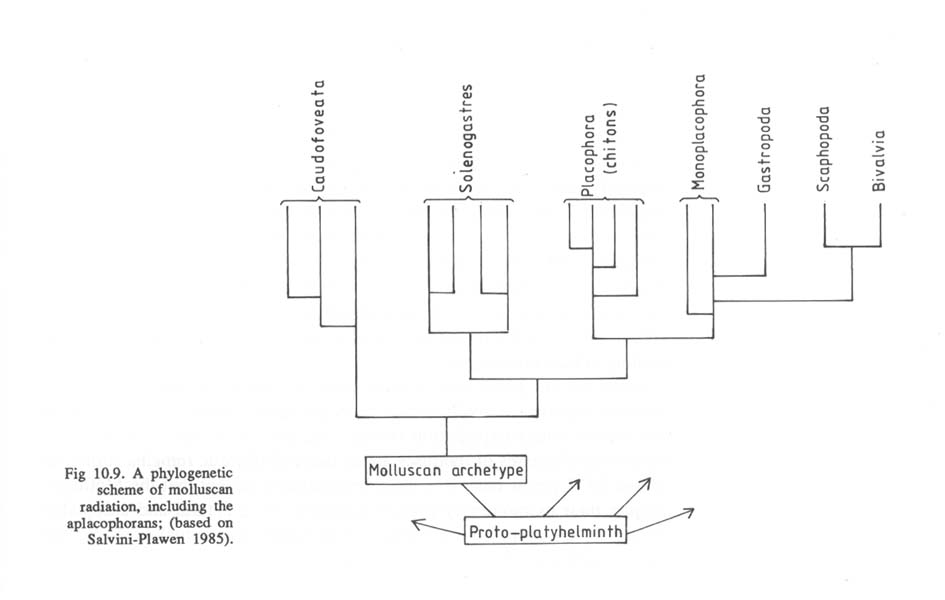
Sieht man sich dazu einige Stammbäume der Mollusken an, so fällt auf, dass auch diese Formen nach den bisher gängigen Hypothesen von Vorfahren ohne Augen abstammen (aus P. Willmer 1990, pp. 266/267):
Soweit ich das bisher übersehen kann, gehen alle Autoren bei solchen Schemata davon aus, dass nicht nur die Ausgangsformen ("Protoplathyhelminthen" und/oder andere), sondern auch die "primitiven Mollusken" ("primitive monoplacophoran-like form") nicht mit Augen ausgestattet waren. Das trifft genauso auf den "Archetyp" der Mollusken zu. Der Grund ist auch unmittelbar verständlich, wenn man sich vergegenwärtigt, dass Augen bei den Niederen Mollusken gar nicht vorkommen und bei den Muscheln (Bivalvia) nicht die Regel, sondern die seltene Ausnahme sind (Land 1981). Auch kann innerhalb der Mollusken kein einheitlicher Stammbaum rekonstruiert werden, nach dem man die verschiedenen in dieser Tiergruppe vorkommenden Augentypen durch gleitende Übergänge voneinander ableiten könnte. Das ist nicht einmal innerhalb der Klasse der Bivalvia möglich (vgl. die vorliegende Schrift, pp. 90-110): "Mit anderen Worten: Das everse Grubenauge von Arca barbata (Arcaceae) hat sich nicht zum eversen Lochkameraauge von Lima squamosa (Pectenaceae) und dieses nicht zum Linsenauge von Tridacna maxima (Cardiaceae) weiterentwickelt". - Zu den paläontologischen, anatomischen und systematischen Details vgl. pp. 90 ff.. Die Abbildung in der Augenschrift 1989, p. 91, vermittelt eine erste Übersicht über das verstreute und seltene Auftreten von Augen bei Muscheln.
Wenn man für die Mollusken einen gemeinsamen mit Augen ausgerüsteten Vorfahren postuliert, dann müsste man auch annehmen, dass zahlreiche Linien diese - selektionstheoretisch allgemein als äußerst vorteilhaft geltende Erfindung - wieder eingebüßt und innerhalb solcher Gruppen (wie der Muscheln) nur in ganz seltenen Fällen bis heute beibehalten und eventuell noch weiterentwickelt hätten. (Man könnte nun eine Detailstudie verfassen, für welche Linien der Mollusken Lichtsinnesorgane zumindest nicht so eindeutig nachteilig wären, dass sie unbedingt wegselektioniert werden müssten. Überdies würde sich die Frage nach rudimentären Augen stellen.)
Mit einem Wort: Auch im Rahmen der Evolutionstheorie sprechen bisher alle paläontologischen (zeitliche Reihenfolge des Auftretens der Formen ohne und mit Augen), anatomischen und systematischen Fakten gegen die Annahme einer mit Augen versehenen Ausgangsform für die Mollusken - mit Ausnahme von Pax-6-homologen Genen und vielleicht (und hoffentlich!) noch mehreren weiteren Regulatorgenen.
Ähnliche Schwierigkeiten ließen sich im Detail auch für weitere Tiergruppen herausarbeiten - wie schon für die Wirbeltiere ansatzweise gezeigt.
Wenn man nun nicht sämtliche paläontologischen und anderen Tatsachen auf den Kopf stellen und damit eine ganze Kette völlig neuer und wohl zumindest vorläufig kaum oder gar nicht lösbarer evolutionstheoretischer Probleme schaffen möchte, wird man wohl nach einer anderen Erklärung für die Verbreitung und Funktion von Pax-6-homologen Genen als der direkten Abstammung aller Pax-6 -Homologen von einem Prototyp suchen müssen.
Dazu seien einige Möglichkeiten aufgeführt:
Möglichkeiten innerhalb der Evolutionstheorie:
1. Mehrfach konvergente Entstehung der Pax-6-ähnlichen Sequenzen. Dieser Ansatz ist jedoch mathematisch derart unwahrscheinlich, dass man ihn eigentlich gar nicht weiter zu diskutieren braucht. Denn die konvergente Entstehung längerer spezifischer DNA-Sequenzen potenziert die ohnehin schon bestehenden Unwahrscheinlichkeiten der Bildung einzelner spezifischer DNA-Sequenzen (vgl. Lönnig 1989) ins Unermessliche.
2. Horizontaler Gentransfer: Diese Erklärungsmöglichkeit - von Syvanen erst neulich wieder recht positiv referiert (Ann. Rev. Genet. 28: 237-261; 1994) - ist jedoch ebenfalls zu unwahrscheinlich, um damit bei höheren Organismen in der Praxis ernsthaft zu rechnen und weit reichende Probleme lösen zu können (Lönnig; Artbegriff pp. 462/463)
Möglichkeiten außerhalb der Evolutionstheorie:
Obwohl in Biologenkreisen heutzutage fast generell und grundsätzlich "verboten" bzw. für viele Zeitgenossen überhaupt nicht mehr als eine Denkmöglichkeit vorhanden, würde die "intelligente DNA-Codierung" (wie ich diesen Ansatz zu nennen pflege) alle mit den heutigen Evolutionstheorien verbundenen Schwierigkeiten mit einem Schlage lösen: Pax-6 , zum Beispiel, und viele andere Sequenzen könnten dann tatsächlich mehrfach "konvergent" entstanden sein, aber die Ursachen wären keine unwahrscheinlichen "Zufalls-Mutationen" (Duplikationen mit einer daran anschließenden hochgradig unwahrscheinlichen Kette von Nucleotid-Substitutionen, die zu Pax-6-ähnlichen Genen führen), sondern die gezielte DNA-Programmierung, wobei bewährte Sequenzen für ähnliche Funktionen für die verschiedensten Tiergruppen verwendet wurden (ein Gedanke, der gentechnologisch versierten Biologen vom Prinzip her eigentlich keinerlei Schwierigkeiten bereiten sollte). Mir erscheint jedenfalls dieser Ansatz am überzeugendsten. (Grundsätzliche Einwände dazu habe ich in der Artbegriffsarbeit 1993 referiert
(Artbegriff, Evolution und Schöpfung) und Falsifikationskriterien habe ich 1991 genannt; vgl. Details unter Kann der Neodarwinismus durch biologische Tatsachen widerlegt werden?).Untermauert wird dieser Ansatz weiter dadurch, dass wir um das Konvergenzproblem letztlich gar nicht herum kommen (siehe Nachtrag 2002). Gehring rechnet damit, dass "more than 2 500 genes" in der Morphogenese des Drosophila-Auges involviert sind (1995, p. 1791) und die Fragestellung von p. 1792 (zukünftige Forschung, Vergleich Drosophila-Facetten-Auge/Maus-Auge:"...how many new genes have been recruited into the developmental pathways in the course of evolution.") möchte ich dahin gehend erweitern, indem ich frage: "How many "old" genes, how many entirely new ones and how many convergently programmed genes have been recruited and programmed into the developmental pathways for the origin of these organs?"
Es gibt zahlreiche weitere gute Gründe (Lönnig 1989, 1991, 1993) anzunehmen, dass für den Ursprung der hochkomplexen synorganisierten Augenstrukturen der Ansatz der intelligenten DNA-Codierung - in Verbindung mit den bekannten Mutationserscheinungen zur Erklärung der neutralen Variation sowie der Degeneration von Organen - letztlich alle Hauptprobleme lösen wird.
NACHTRAG 2002
Fast alles ist zur Konvergenzfrage noch im Fluss: Es gibt Beispiele für das Vorhandensein von Pax-6-Sequenzen im Tierreich ohne Augenbildung. Die Linsenaugenbildung müsste sowieso 15mal unabhängig voneinander abgelaufen sein, wenn man nicht schon einen recht komplexen gemeinsamen Vorfahren mit Linsenaugen annehmen möchte etc. Um die Konvergenzfrage zu vermeiden, müsste man im letzteren Fall auch sämtliche einfacheren Augenbildungen im Tierreich von Linsenaugen ableiten, was zu vielen weiteren evolutionstheoretischen Problemen führen würde. - Es wäre sehr gut und empfehlenswert, wenn sich ein oder mehrere Intelligent-Design-Theoretiker mit den notwendigen zeitlichen und wissenschaftlichen Möglichkeiten mit diesen Evolutionsfragen weiter gründlich-kritisch auseinander setzen könnte(n)!
Intelligent Design ist die beste wissenschaftliche Erklärung für den Ursprung der synorganisierten Augenstrukturen (vgl. weiter
Die Synthetische Evolutionstheorie und die Intelligent Design-Theorie: Ein Vergleich).