INSELPOPULATIONEN
Wiederholt haben wir in unserer Arbeit schon Degenerationserscheinungen bei Inselpopulationen angesprochen, einen Punkt, den ich mit dem Hinweis auf einige generelle Trends noch etwas erweitern möchte. Williamson behandelt im zweiten Kapitel seiner Arbeit ISLAND POPULATIONS (1981), betitelt "Features of island life" nach historischen Rückblick auf die grundlegenden Erkenntnisse Hookers (um 1866) folgende Unterthemen (Numerierung von mir):
(1) The biota of Tristan da Cunha
(als Musterbeispiel für die generellen Trends zu:)
(2) Impoverishment
(...fewer species on an oceanic island than on the same-sized area of a continent.)
(3) Disharmony
(Ungewöhnliche Ökosysteme mit veränderten Nahrungsketten; auf Tristan z.B. keine Raubvögel, die dort wohl auch nicht als Population existieren könnten; generell keine Landsäugetiere und Amphibien, was offensichtlich eine Frage der Verbreitungsmöglichkeiten ist. Die Populationen sind verwundbarer; Eindringlinge fremder Arten haben größere Chancen, sich durchzusetzen als auf Kontinentalgebieten; die Spezies neigen dazu, schneller als ihre kontinentalen Verwandten auszusterben.)
(4) Dispersal
(Gute Verbreitungsmöglichkeiten sind Voraussetzung, auf ozeanische Inseln zu gelangen.)
(5) Loss of dispersal ability
(Einige Punkte haben wir mit den obigen Zitaten schon belegt. Williamson führt weitere Beispiele aus dem Tier- und Pflanzenreich auf. Nach Hinweis, dass man bei der Interpretation flügelloser Insekten und flugunfähiger Vögel vorsichtig sein sollte, da solche Formen auch auf den Kontinenten weit verbreitet sind, lesen wir p. 46:
"Nevertheless, island environments frequently seem to encourage the evolution of the loss of dispersal ability. Two reasons are often put forward for this loss. The first is that with the absence of predators, flying no longer has a selective advantage, and indeed might have a disadvantage in that the energy used in flying could be more usefully put to other purposes. The second explanation is that flying forms are more likely to be swept to sea by high winds, giving a selective advantage to the flightless ones. Both explanations could well be true, but there is a lack of hard evidence."
Hinzuzufügen ist noch, dass der auf abgelegenen Inseln u. U. verhängnisvolle 'Wandertrieb' nach Loss of dispersal ability nicht mehr in die Tat umgesetz werden kann.)
Die Anhäufung von unter bestimmten Umständen neutralen und schwach nachteiligen Allelen ist nach den oben (pp. 340, 358) zitierten Mutationsraten auch ganz ohne Selektion möglich. Es ist gerade der Ausfall des 'Selektionsdrucks' (Tiere, die sonst von Raubtieren gefressen werden und die durch Funktionsbeeinträchtigung verschiedener Organe normalerweise ihre Umwelt gar nicht mehr bewältigen könnten, können sich auf Inseln oft noch behaupten), der die Degeneration verschiedener Funktionen erst in größerem Maßstab ermöglicht. Abgelegene Inseln (im symbolischen Sinne auch auf den Kontinenten) bieten somit für einen solchen Prozess die besten Voraussetzungen. Auch an dieser Stelle sei wieder betont, dass Selektionsprozesse für Inselpopulationen mit diesen Überlegungen nicht völlig ausgeschlossen werden sollen, aber es handelt sich bei der Selektion nur um einen Faktor unter vielen anderen.
Williamson behandelt im zitierten Kapitel desweiteren folgende Unterthemen:
(6) Reproductive change
(Anhand der Entengattung Anas zeigt der Verfasser, dass zwar das Gewicht der Eier einigermaßen konstant ist, dass aber wegen des häufiger festgestellten geringeren Körpergewichts der Inselformen das Gewicht der Eier in Relation zum Körpergewicht zunimmt.)
(7) Size change
(p. 48:) "Island endemics may be markedly larger, or appreciably smaller, than their mainland relatives."
(8) Extinction
(Von Menschenhand verursachtes und 'natürliches' Aussterben.)
(9) Relicts
(Formen, die einst weit verbreitet waren und sich nur auf Inseln erhalten haben.)
(10) Endemics
(Rekombinanten, die als Populationen nur auf Inseln nachgewiesen worden sind.)
Die Tendenz zur Degeneration ist bei Inselpopulationen unverkennbar.
So haben die vielzitierten Darwinfinken ein vergleichsweise kleines Herzgewicht, "bei der schwersten Art mit 5,4 v.H. des Körpergewichts kleiner als das fast aller anderen Vögel". Vom wendigen und lebhaften Laubsängerfinken abgesehen, wirkt der Flug der Darwinfinken schwerfällig. "Aber auch die plump schwirrenden Arten finden ihren Weg durch das Gezweig, mit dem die Galapagosinseln reich gesegnet sind" (Curio 1980, pp. 344/345).
Von den symbolischen Inseln auf den Kontinenten hatten wir schon (p. 114) die Höhlen mit ihren Bewohnern genannt und die Arbeit Culvers (1982) zitiert. Schemmel, Parzefall, Peters und Peters und andere haben zu diesem Thema weitere informationsreiche Beiträge 1984 geliefert. Im Vordergrund stehen Strukturabbauphänomene.
An dieser Stelle könnte vielleicht der Einwand erhoben werden, dass bei manchen Beispielen von Inseln- und Höhlenbewohnern, ja überhaupt beim Thema "regressive Evolution" nicht nur Informations-, Struktur- und Funktionsverluste zu verzeichnen sind, sondern auch das vermehrte Auftreten bestimmter Strukturen auffällt. So ist bei der Höhlenfischform von Astyanax mexicanus die Anzahl und Verbreitung der Geschmacksknospen im Vergleich zur Flussfischform stark erweitert. "Der oberirdisch lebende Flußfisch erfaßt seine Beute optisch und prüft sie auf ihre Genießbarkeit durch Geschmacksknospen in der Mundhöhle. Das Höhlenderivat findet seine Beute durch kreisendes Absuchen des Bodens mit Lippen und Kopfunterseite. Es ist keine Frage, daß hierfür Zahl und Ausbreitung der Geschmacksknospen von Bedeutung sind, wie an vielen bodenlebenden Fischarten zu beobachten ist Demzufolge sind bei Anoptichthys solche Gene schrittweise angereichert worden, die zur Vermehrung der Knospenzahl und zur Ausbreitung des geschmacksrezipierenden Areals in der äußeren Kopfhaut beitragen" Schemmel 1984, p. 13. Die Deutung der Rekombination in einem polygenen System erscheint überzeugend (vgl. Rekombinationsquadrat p. 80). Man möchte jedoch eher an Regulatorgene als an Strukturgene in diesem Zusammenhang denken, denn an dem Aufbau der Geschmacksknospen selbst scheint sich nichts verändert zu haben. Die Erweiterung des Geschmacksfeldes könnte damit auch in wenigen Schritten erreicht worden sein. Wie dem auch sei, das genetische Potential für die Anreicherung der Geschmacksknospen war offenbar von vornherein gegeben und ist hier auch mehrfach voneinander in verschiedenen Populationen realisiert und für brauchbar befunden worden.
Es ist dabei folgende Deutung möglich:
Die Stammform beider Linien realisierte das umfassende genetische Potential sowohl für die vollständige Augenausbildung als auch das große Geschmacksknospenfeld. Bei den Höhlenbewohnern degenerierte das Potential für das erstere, bei den Flussbewohnern das für das letztere. Vorsichtiger formuliert sind geschmacksknospenreiche Rekombinanten l000de von Malen schon bei der Flussform aufgetreten und haben sich dann bei der Höhlenform als besonders zweckmäßig erwiesen. Von der Entwicklung völlig neuer Strukturen und Systeme kann nicht die Rede sein. Die Höhlenformen waren 'präadaptiert'. Schemmel zeigt das sehr deutlich für die Raumorientierung bei Astyanax (p. 14):
Der oberirdische Flußfisch besitzt große, stäbchenreiche Augen, die ihn als Dämmerungstier auszeichnen. Für diese Lebensweise wird die Orientierungsfunktion des Auges noch zusätzlich durch ein weiteres Sinnesorgan, das dem Ferntastsinn dient, unterstützt. Es handelt sich um das Seitenliniensystem, das für alle niederen im Wasser lebenden Vertebraten bezeichnend ist. Die als Neuromasten benannten Sinneshügel besitzen Sinneshaare, die von einem elastisch beweglichen Gallerthügel umschlossen werden. Geringste Bewegungen und Staudruckerscheinungen des Wassens stellen die biologisch adäquaten Reize für diese Strömungsrezeptoren dar, die in Kanälen versenkt oder frei in der Epidermis sitzen.
Beim Flußfisch sind vor allem die freien Neuromasten in einer Vielzahl entwickelt, wie sie bei anderen Fischgruppen nicht zu finden sind..........Der Flußfisch kann sich deshalb auch ohne Augen recht gut orientieren und ist nicht zur Bewegungslosigkeit verurteilt. Im Gegenteil, geblendet tastet er gleichsam durch ständige Schwimmbewegungen Raum und Hindernisse ab. Dies bedeutet, daß Flußfische aufgrund ihrer Lebensweise zwei verschiedene Organe für die gleiche allgemeine Funktion besitzen. Die Höhlencharaciniden übernahmen das hochentwickelte Lateralsystem der Flußfische. Das Überleben in unterirdischen Biotopen wurde dadurch gesichert, daß der Ferntastsinn in der Lage war, den Gesichtssinn zu ersetzen. Durch Funktionsträgerwechsel wurde die Entwicklung zum Höhlenfisch eröffnet.
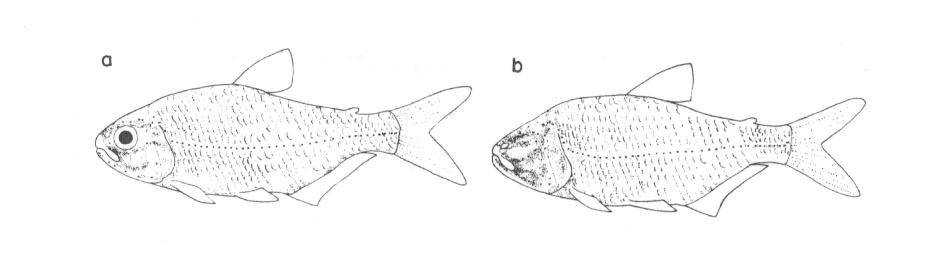
Abb. 55: Dichte und Verbreitung epidermaler Mechanorezeptoren beim Flußfisch Astyanax mexicanus (a) und seinem Höhlenderivat (b). Das hochentwickelte Neuromastensystem ermöglicht auch dem Flußfisch schon die räumliche Orientierung im Dunkeln. Aus Schemmel 1967/1984.
Jaennel hat 1923 festgestellt, dass für viele Coleopteren die unterirdische Umwelt der Höhlenbewohner mehr ein konservierendes als ein modifizierendes Medium ist. "Nach JEANNEL sind z.B. die langen Tasthaare auf den Elytren beim cavernicolen Aphaenops ein ursprüngliches Merkmal der Trechini, das bei den oberirdisch lebenden Formen sekundär reduziert wurde. Auch LELEUP (1956) vergleicht die anophthalme und depigmentierte humicole Fauna der tropischen Gebirgswälder Zentralafrikas mit der cavernicolen Fauna Nordamerikas und Europas und bemerkt aufgrund der Ähnlichkeit, daß die Vorfahren der europäischen Cavernicolen ebenso im Humus großer feuchter Gebirgswälder des Tertiärs gelebt haben müssen wie heute noch die afrikanische Fauna" Schemmel, p. 21. Vandel hat 1965 die Höhlenformen der Tierwelt als Beispiele phyletischer Seneszenz interpretiert.
Höhlen- wie auch Inselformen dürften häufig einen anderen Bereich des ursprünglich umfassenderen genetischen Potentials aufrechterhalten haben als ihre nicht-höhlenbewohnenden bzw. kontinentalen Verwandten. Dieser Punkt wird in vielen Abhandlungen wie der 'Kompensation in der Höhlentierentwicklung' (Schemmel) nicht genügend berücksichtigt.
Weiter sei erwähnt, dass bereits der Ausfall bestimmter Strukturen in dem fein ausbalancierten physiologischen Getriebe zur Anreicherung von Stoffwechselprodukten mit Rückwirkung auf die Entwicklung des Gesamtsystems verbunden sein kann. Entsteht zum Beispiel ein genetischer Block in einer Genwirkkette, so kommt es zur Anreicherung des oft schwer weiter umsetzbaren Zwischenprodukts. Die Phynylketonune beim Menschen kann als Veranschaulichung dienen: Dadurch, dass bei den Betroffenen die Umwandlung von Phenylalanin zu Tyrosin weitgehend unterbleibt (Defekt in der Phenylalanin-4-Hydroxylase), kommt es zu einer Anreicherung von Phenylalanin im Blutserum, im Harn und im Liquor. "Durch die starke Anreicherung von Phenylalanin werden Nebenwege beschritten, die zur Bildung von Phenylbrenztraubensäure, Phenylethylamin, Serotonin u.a. führen. Wahrscheinlich schädigen einige dieser Nebenprodukte das Gehirn. Die Kranken sind geistig oft stark gestört, meistens bis zur Idiotie" (Günther 1984, p. 185).
Wenn auch häufig nicht in vergleichbar krasser Form, so dürfte Funktionsausfall vieler Gene mit mehr oder weniger additiven Rückwirkungen auf das Gesamtsystem verbunden sein, so dass sich die Unterschiede zwischen unterschiedlich degenerierten Populationen einer Art aufgrund solcher Rückwirkungen noch weiter verstärken.
Auf der anderen Seite könnte der Ausfall eines Systems mit der Erhöhung der Freiheitsgrade in der Expression eines zweiten Systems einhergehen (bei Kulturpflanzen scheint das desöfteren der Fall zu sein), solange jedenfalls die oben zitierten Rückwirkungen hier keinen Strich durch die Rechnung machen.
Zu jedem der oben nach Williamson zitierten Punkte ließe sich eine Spezialarbeit anfertigen, was genauso für viele weitere der in diesem Kapitel zitierten Beispiele gilt. Ich möchte im Rahmen dieser Arbeit nur feststellen, dass sich die meisten Besonderheiten von Insel- und Höhlenpopulationen ganz mühelos dem Degenerationsthema zuordnen lassen.
Völlig neue Formen entstehen in keinem Falle. Viele der von Systematikern als eigene Arten und Gattungen beschriebenen Formen sind miteinander kreuzbar und können zum Teil noch fruchtbare Nachkommen miteinander hervorbringen (wie z.B. die Fluss- und Höhlenform von Astyanax mexicanus, bei der die Höhlenform zunächst sogar als eigene Gattung (Anoptichthys) beschrieben worden war, die 'Arten' der Gattung Anas (vgl. pp. 27, 197) und viele Drosophilaarten (vgl. p. 21).)
Zwar sind durch Degeneration zahlreiche sekundäre Sterilitätsbarrieren entstanden, aber nach dem genetisch-plasmatischen Artbegriff keine neuen Arten (pp. 122-137, 210-283).
Als eindrucksvolle Evolutionsbeweise liest man desöfteren, dass auf manchen ozeanischen Inseln nur endemische Arten leben. Fragt man nach der Zahl der endemischen Gattungen, so nimmt der Prozentsatz schon stark ab und bei der Frage nach dem Anteil endemischer Familien ist er in der Regel auf Null gesunken. Dabei muss man sich noch bewusst machen, dass auch der Familienbegriff starken Schwankungen unterworfen sein kann (vgl. pp. 36, 57, 276). Was an eindrucksvollen Zahlen endemischer Arten vorgelegt wird, sind fast ausschließlich Arten im Sinne des morphologischen Artbegriffs (vgl. pp. 61-66). Dasselbe trifft auf die Mehrzahl der Gattungen zu. Die Frage nach dem genetischen Artbegriff wird in diesem Zusammenhang selten gestellt.
Sehen wir uns dazu ein Beispiel näher an. Ayala legte 1977, p. 273 eine Tabelle der endemischen Flora und Fauna von Hawaii vor. Lobenswerterweise werden hier wenigstens noch die Prozentsätze der Gattungen mitaufgeführt. Ich habe die Tabelle im folgenden wiedergegeben und den Prozentsatz der nicht-endemischen Genera zur besseren Veranschaulichung aufgeführt und im Schriftbild hervorgehoben:
Tabelle 15 (aus Ayala 1977):
|
|
Genera (Total number) |
Percent endemic |
Percent nonendemic |
Species, subspecies, varieties |
Percent endemic |
|
Ferns |
37 |
8% |
92% |
168 |
65% |
|
Flowering plants |
216 |
13% |
87% |
1729 |
94% |
|
Land molluscs |
37 |
51% |
49% |
1064 |
99+% |
|
Insects |
377 |
53% |
47% |
3750 |
99+% |
|
Drosophilids |
4 |
75% |
25% |
510 |
100% |
|
Birds |
41 |
39% |
61% |
71 |
99% |
Wenn nur 30 heterozygote Allelpaare* bereits 1 073 741 824 morphologische Varianten, Subspezies und Spezies ermöglichen können und 50 solcher Genpaare 1 125 890 000 000 000 (d.h. über 1 000 Billionen) Rekombinanten zulassen, dann ist man fast erstaunt, dass bei den Spezies, Subspezies und Varietäten der Farne und Blütenpflanzen der Anteil der endemischen Formen nicht höher ist. Denn weder bei einer Zufallsverteilung noch unter der Annahme der Selektion auf allen Ebenen könnte man bei einem solchen Rekombinantenpotential einen größeren Prozentsatz identischer bzw. sehr ähnlicher Linien und Spezies bei längerer Trennung von Arealen erwarten (die Hauptinseln Hawaiis umfassen immerhin eine Fläche von mehr als 16 600 km2, Pearl Harbor ist ca. 3 870 km von San Francisco entfernt. Die Insel Hawaii wird auf 500.000 Jahre und Kauai auf 5,6 Millionen Jahre datiert (zunehmendes Alter nach Nordwesten). Weitere inzwischen versunkene Inseln werden bis in das späte Mesozoikum zurückdatiert.
Da die Gattungen zum großen Teil ebenfalls ohne Rücksicht auf die Genetik rein morphologisch definiert sind und zum Teil schon durch den Artbegriff der Synthetischen Evolutionstheorie starke Revisionen erfahren haben - von 6 000 bis 7 000 um 1920 auf derzeit etwa 1 700 bis 1 800 bei den Vögeln (vgl. p. 49), und dabei beachte man die Abwehrhaltung Mayrs gegen den genetischen Artbegriff (zitiert p. 197) - ist selbst noch die Zahl der endemischen Gattungen relativ gering. Die Zahl endemischer Familien wird in evolutionistischen Arbeiten in der Regel nicht angegeben. Shute fand 1961, p. 155 nach Überblick über die Besonderheiten ozeanischer Inselpopulationen, das weltweit nur vier Familien bis dahin nachgewiesen worden waren, die ausschließlich auf Inseln und lange isolierten Binnengewässern vorkamen. Er schreibt:
...distinct families are found only on Madagascar, Hawaii and New Zealand, and in Lake Baikal, an equally long isolated sea. Even such family formation is rare, amounting to only four examples, which show how exceptional such an event has been.
Dewar schrieb 1957, p. 162 zur Frage nach der Evolution auf ozeanischen Inseln:
Many of these Islands have been in existence for a very long time, yet no new family of animals has evolved on any of them, all the animals which inhabit them belong to mainland families. The nearest approach to peculiar families on a true oceanic island are the sub-family of flowerpeckers, Drepaninae and the sub-family of land snails the Achatinellae of the Sandwich Islands.
...Ex hypothesi oceanic islands have been peopled by immigrants from the continents, and, as they were originally uninhabited, they are the places at which Evolution should be most rapid. The environment would be different from that of the mainland, and for a time there would be no enemies to fear and little competition for food, so that variation could have free play and not be subjected to the pruning hook of natural selection. Thus the fact that no new family has evolved on an oceanic island suggests that it is not possible to become converted into a different one.
Wie auf p. 276 gezeigt, kann der Familienbegriff im konkreten Falle von Autor zu Autor unterschiedlich gebraucht werden und verschieden große Tier- und Pflanzengruppen umfassen. Heute findet man in den meisten Evolutionistischen Arbeiten z.B. die Drepaninae als eigene Familie (Drepanididae) aufgeführt. Bock kommentiert die Frage nach dem Ursprung und den Verwandtschaftsverhältnissen dieser Vogel-(Sub)-Familie 1970, p. 707:
The origin of the Drepanididae and the precise relationships of the Drepanididae and the Psittirostrinae are still largely unresolved. Disagreement still exists as to whether the drepanidids evoved from a nectar-insect feeding stock (Thraupidae-"Coerebidae" complex) or from a seed-insect feeding finch (Fringilla-like member of the Carduelinae) but the evidence is not yet available. Some doubt also exists as to whether the Drepanidinae and Psittirostrinae evolved monophyletically from a common ancestor on the Hawaiian Island although no real evidence for a possible diphyletic origin of the Hawaiian honeycreepers has been presented.
Darüber hinaus besteht noch die Möglichkeit, dass es sich bei diesen Vogelgruppen um Reliktformen handelt, die einst ein wesentlich größeres Areal besiedelten und - wie die Tuatara auf Neuseeland - heute nur noch auf einer isolierten Inselgruppe vorkommen. Die Drepaniden zeigen übrigens an mehreren Arten typische Kennzeichen der Degeneration wie Flugunfähigkeit (Loss of dispersal ability) und die Tendenz, schneller auszusterben als kontinentale Vogelgruppen. Austin schreibt schon 1961/1963, p. 284:
Die Kleidervögel bieten ein eindringliches Beispiel für die Tatsache, dass sich eine allzu schnelle Entwicklung und Überspezialisation ungünstig auf die Lebensfähigkeit einer Art auswirkt. Von den 22 bekannten Arten sind 9 bereits ausgestorben und weitere 8 oder 10 vegetieren nur noch in kleinen, örtlich begrenzten Populationen. Verschiedene Arten, die früher weit verbreitet über das ganze Inselgebiet lebten, trifft man nur noch auf 1 oder 2 Eilanden an. Die heute noch am häufigsten vertretenen Arten stellen auch die am wenigsten spezialisierten dar.
Willis stellte im Rahmen seiner pflanzengeographischen Studien 1949, p. 159 fest, dass nahezu alle endemischen Arten Hawiis weitverbreiteten (und alten) Familien und Gattungen angehören.
Auf dem im Pazifik gelegenen Neukaledonien sind die Phanerogamen mit 108 Familien, 365 Genera und 1511 Spezies vertreten. Der Anteil der endemischen Formen beträgt (in derselben Reihenfolge): 4 %, 22,4 % und 89,9 % (Radovsky et al. 1984).
Die im Indischen Ozean gelegenen (und auf 150 Millionen Jahre datierten) Seychellen weisen neben einer endemischen Blindwühlenfamilie nur eine Froschgruppe mit unsicherer Verwandtschaft auf (die Sooglossiden [nach Heusser noch die Unterfamilie der Sooglossinae (Grzimeks Tierleben Bd. 5, 1980, p. 404)]) sowie eine ebenfalls in den Familienrang erhobene monotypische Pflanzengruppe, die Medusagynaceae (Stoddart 1984).
Freytag schreibt über die Blindwühlen (Gymnophiona) 1980, p. 356, dass bisher alle Formen in einer Familie zusammengefasst wurden, dass jedoch neuerdings von Taylor drei Familien unterschieden werden (vgl. das Haeckel-Zitat p. 35).
In diesem Zusammenhang sei noch einmal an die oft merkwürdige Verbreitung von Reliktformen erinnert. So kommt z.B. der Farn Thrysopteris nur in Japan und auf den Juan Fernandez Inseln (Chile) vor. Würde man diesen Farn nur von den Juan Fernandez Inseln kennen, so könnte man leicht auf den Gedanken kommen, der Farn hätte sich dort 'entwickelt' und wäre sonst nirgends aufgetreten. So aber deutet man diesen Fall als Reliktform (Steussy 1984).
Mehrere Autoren haben übrigens auf die wiederholt festgestellte mangelnde Korrelation zwischen geographischer und genetischer Diversität hingewiesen. So schreiben Narsinghani et al. in ihrer Arbeit GENETIC DIVERGENCE IN PEAS 1978, p. 378:
The earlier workers assumed that geographical diversity reflected genetic diversity. However, Moll, Salhuana and Robinson (1962), Timothy (1963) and Murthy and Arunachalam (1966) could not find any direct relationship between geo-graphic distribution and genetic divergence. In this study also, the clustering pattern indicated that genotypes from different origins had clusters together in five out of eight cases. This further supports the view that geographical and genetic diversity may not be related.
Diese Frage ist u.a. für die Züchtungsforschung von unmittelbarer Bedeutung. Singh et al. sind zu dem gleichen Ergebnis bei ihren Untersuchungen der genetischen Divergenz bei Pennisetum gekommen und berichten 1981, p. 187 u.a.:
Genetic diversity has been found to be associated with geographic diversity by Joshi and Dhawan (1966) and Ram and Panwar (1970), but Murthy and Arunachalam (1966) and Singh and Gupta (1968) have shown that geographical distribution and genetic diversity need not be directly related. The present findings indicated that no firm conclusion regarding relation between genetic divergence and geographical distance, more particularly in the population under study, could be made as a few parents of different places of origin fall in the same cluster while parents of the same place were distributed into different clusters.
Zudem findet sich desöfteren auch keine klare Beziehung zwischen der geographisch-morphologischen und der genetischen Distanz. Second untersuchte 181 Linien der Oryza sativa-Gruppe an 24 Isozymloci und bemerkt 1984, p. 45:
It was a surprise that the morphologically divergent American strains could not be distinguished clearly from their Asian counterparts.
Zu ähnlichen Feststellungen sind Jain et lL. 1980 beim Fuchsschwanz gekommen (LEVFLS OF MORPHOLOGICAL AND ALLOZYME VARIATION IN INDIAN AMARANTH: A STRIKING CONTRAST) und Edwards 1980 bei den Wildformen der australischen Baumwolle (Gossypium australe).
Weiter kann die morphologische Ähnlichkeit bei weit voneinander entfernten Populationen größer sein als bei nahe zusammenlebenden Formen einer Art.
Tabachnik und Powell heben in ihrem Beitrag A WORLD-WIDE SURVEY OF GENETIC VARIATION IN THE YELLOW FEVER MOSQUITO Aedes aegypti 1979, p. 225 z.B. hervor:
Our results show a strong corellation between electrophoretic variation among populations with similar behavioural, morphological and ecological differentiation, regardless of geographic proximity. For example, domestic A. ae. aegypti in East Africa show greater genetic relatedness to domestic populations in the New World than they do to sylvan populations less than a kilometre away. On the other hand, West African A. ae. formosus show greater genetic similarity to East African sylvan populations than to all other domestic populations throughout the world.
Die Autoren meinen, dass Habitat-Selektion den Schlüssel zur Erklärung liefert. Der letzte Punkt des obigen Zitats relativiert allerdings diese Position. Immerhin findet sich hier bei einigen Populationen die eigentlich als Regel erwartete Korrelation elektrophoretischer, ethologischer, morphologischer und ökologischer Daten, - wenn auch auf relativ schmaler Basis und unabhängig von der geographischen Distanz. Zu den Schwachpunkten der Korrelation bemerken die Autoren auf der Seite 226 u.a.:
The genetic distance values (D) detected between subspecies of A. aegypti are small when compared to similar values obtained in species of Drosophila (Ayala, 1975). The relatively small distances are expected since the genetic differentiation among populations is the result of allelic frequencies and not allelic substitutions. No single locus is 'diagnostic' for either subspecies or geographic area. Apparently, in spite of large morphological and behavioural differences, protein Evolution in A. aegypti has not been rapid.
Ich möchte an dieser Stelle meine Ausführungen zum Thema Inselpopulationen und mangelnder Korrelation von geographischer und genetischer Distanz unterbrechen, obwohl man noch eine Fülle interessanter Details aufführen und diskutieren könnte. Aber das wäre Stoff für eine Spezialarbeit für sich.
(Vgl. auch pp. 112-120 und 186 f.)
Vielleicht hat sich mancher Leser schon gefragt, warum auf ozeanischen Inseln und anderen relativ isolierten Gebieten soviele 'endemische Arten' gefunden werden, wenn eine vergleichbare Formbildung zum Beispiel bei Drosophila melanogaster durch Mutationszüchtung doch offensichtlich nicht möglich ist. Antwort: Bei den 'Arealarten' ist das ursprünglich weit umfangreichere genetische Potential durch Anreicherung mutierter Allele, wiederholten Chromosomenstückverlusten und möglicherweise auch durch die Einschränkung der Genexpression auf plasmatischer Ebene (vgl. pp. 237, 280) in den verschiedensten Gebieten der Erde so weit abgebaut, dass die einstigen Formbildungsmöglichkeiten heutzutage weitgehend verloren sind (bzw. nur noch auf niedrigeren Stufen wie bei der Haustierbildung fortgesetzt werden können).
Die Regel der Rekurrenten Variation gilt selbstverständlich nur für den Teil des genetischen Potentials, der nicht schon unwiderruflich verlorengegangen ist (vgl. p. 358). Auf der anderen Seite dürfte jedoch auch klar sein, dass viele der Formen, die heute nur noch an isolierten Arealen anzutreffen sind, im Zuge der Freisetzung und zunehmenden Degeneration des ursprünglichen genetischen Potentials auch schon wiederholt an anderen Stellen der Erde aufgetreten sind.
____________________
*Vgl. die zweite Fußnote zum Gesetz der rekurrenten Variation.