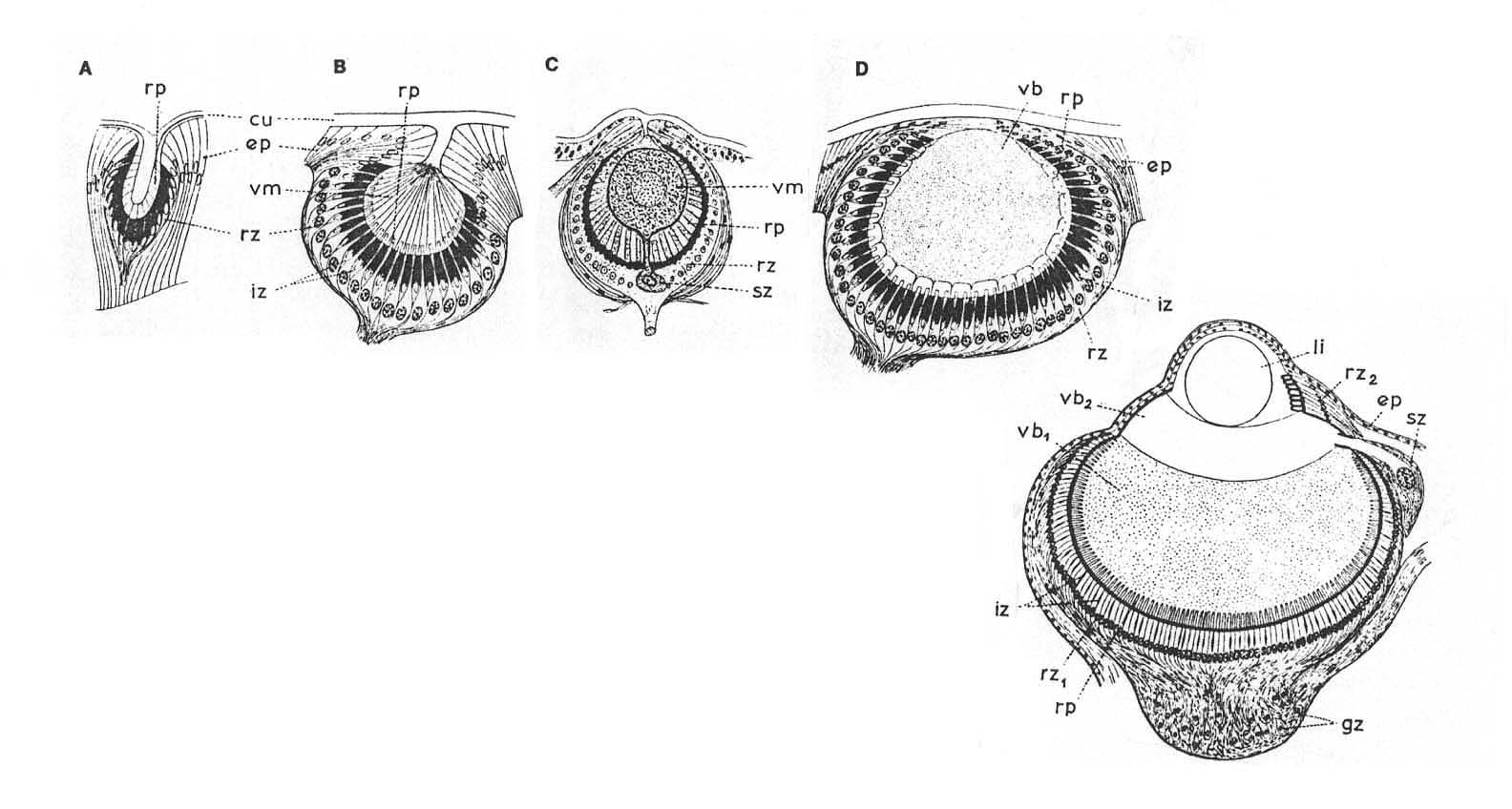
III. Morphologische Serie bei den Polychaeta:
Morphological sequence of differentiation of everse cephalic eyes in Polychaeta (after Hesse, 1899, from Plate, 1924): A, eye pit of Ranzania sagittaria; B, eye cup of Syllis aurantiaca; C, pinhole eye of Phyllodoce laminosa; D, closed eye of Nereis cultrifera; E, lens eye of Vanadis formosa. (cu, cuticule, ep, epidermis: gz, ganglion cells; iz, supportive cells; li, lens; rp, receptoral portion of rz (retinal cells); sz, secretory cell; vb, vitreous body; vm, vitreous mass.)
(Aus Salvini-Plawen und Mayr, p. 240)
Im Text dazu heißt es, dass die Photorezeptoren der Eunicemorpha manchmal mit einer Linse versorgt und hochdifferenziert sind (worauf der Hinweis auf die obige Abbildung folgt), und das besonders bei den pelagischen Alciopidae, die auch eine akzessorische laterale Retina besitzen.
Land bemerkt 1981, p. 514:
Large eyes with distinct lenses have probably only evolved once in this phylum, in the Alciopidae, a family of carnivorous pelagic polychaetes.
Die meisten der auf p. 98 aufgeführten und zuvor im Detail diskutierten Einwände treffen hier genauso zu (siehe dort).
Weiter lässt sich auch bei dieser diskontinuierlichen Sequenz die Serie umkehren: Degeneration durch Mutation an lichtärmeren Biotopen (hier könnte man sogar die Selektion einsetzen) oder durch "neutrale Evolution" ist wesentlich wahrscheinlicher als der Aufbau solcher komplexen Strukturen. Die Serie sieht dann wie folgt aus:
- was sie jedoch aufgrund der verschiedenen nicht voneinander abstammenden Gattungen ebenfalls nicht zur Abstammungsserie machen würde.
Zum Begriff der morphologischen Reihe scheint mir an dieser Stelle ein weiteres Beispiel und eine grundsätzliche Bemerkung angebracht:
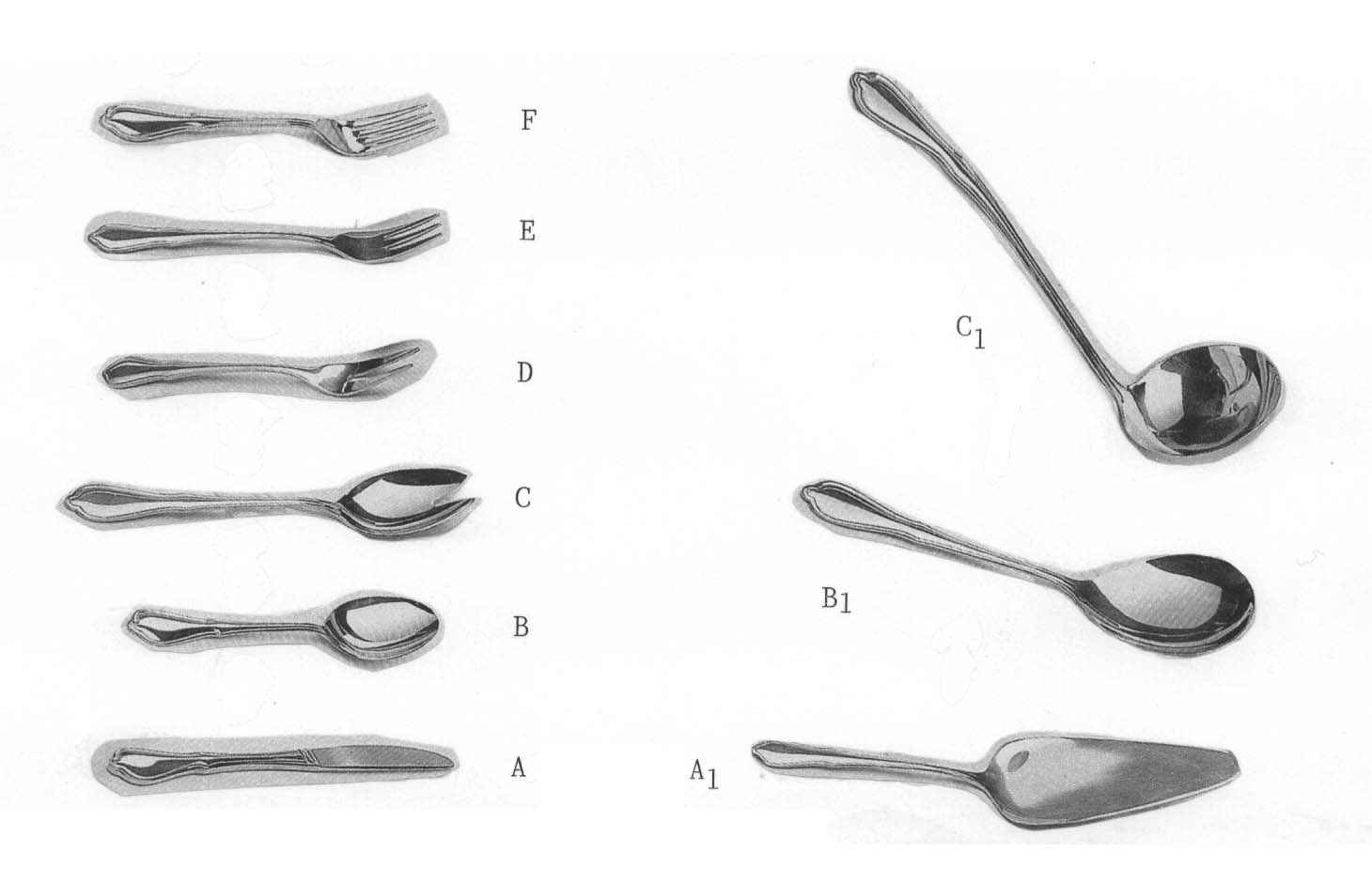
Morphologische Serie beim Besteck: Ableitung der Gabel vom Messer über den Löffel und Sonderentwicklung der Suppenkelle über den Tortenheber. Man beachte besonders die schrittweise Vervollkommnung in der Gabelentwicklung von der 2-zinkigen Fleischgabel (D) über die 3-zinkige Kuchengabel (E) zur 4-zinkigen Essgabel (F). Das Salatbesteck (C) ist das Bindeglied zwischen Löffel (B) und Fleischgabel (D) (Mosaikevolution!). Man braucht nur vorauszusetzen, dass sich alles vom primitiven Messer ableitet.
Wie die Abbildung von p. 102 unten lehrt, kann selbst eine problemlose Abstufung, d.h. ohne größere Diskontinuitäten durch starke Komplexitätszunahme innerhalb der Serie, allein noch nicht die realgenetische Abstammung beweisen. Alle nur denkbaren Formenmannigfaltigkeiten wie Schneekristalle, geometrische Figuren, Musikinstrumente, Schreibmaschinen, Flugzeuge etc. kann man in kontinuierliche Serien mit steigender Komplexität anordnen (worauf im Prinzip schon Galton hingewiesen hat) und mit der Anordnung die Selbstorganisation der Formenreihe durch rein physikalische Gesetzmäßigkeiten über die Zwischenstufen postulieren. Es sind jedoch die Transformationen selbst, die bewiesen werden müssen. Dass Organismen dynamische Systeme sind und sich fortpflanzen - ein häufig an dieser Stelle erhobener Einwand - ändert an der Sachlage nichts (siehe auch pp. 42/43). Solange die postulierten Aufbau-, Evolutions- und Transformationsprozesse durch Mutation und Selektion nicht nachgewiesen werden können, liegt die Evolutionsmethodik bei den eben genannten Beispielen und den Organismen logisch auf der gleichen Ebene.
Übrigens ist auch in der Technik der Zerfall (die "Selbstdesorganisation") allein nach physikalischen Gesetzen der Realität wesentlich näher als der Selbstaufbau.
Mit diesen Ausführungen sei hervorgehoben, dass weder die Anordnung der verschiedenen Augentypen in Richtung steigender Komplexität noch als Serie abnehmender Differenzierungsstufen allein beweiskräftig für einen bestimmten Entstehungsmodus und ihrer Abstammung voneinander sein kann. Realgenetische und reproduzierbare Abstammung bei solchen Serien kennt man jedoch bisher nur in Richtung abnehmender Komplexität: "...wir kennen keine echte Differenzierungsmutation weder in einzelnen Genen noch in einer Kombination von Erbfaktoren" (Remane/Storch/Welsch, vgl. Zitat p. 43).
Wenn wir jetzt zu unserer Frage zurückkehren, ob wir mit einer Serie abnehmender Differenzierungsstufen und Verlustmutationen bereits alle Probleme, die mit den verschiedenen Augentypen verbunden sind, erklärt haben, dann dürfen wir nicht den gleichen Fehler machen wie die Neodarwinisten, nur mit dem isolierten Organ arbeiten und dabei das Gesamtsystem (den Organismus und seine Umwelt) vernachlässigen. Zur Veranschaulichung auf die Besteckserie übertragen: indem wir Zweck und Funktion ausklammern.
Im Laufe der Arbeit wurde schon wiederholt darauf hingewiesen, dass die Organismen zur Bewältigung ihrer Umwelt bereits optimal mit Sinnesorganen ausgestattet sind (vgl. pp. 33, 34, 45, 47, 49, 55-57, 64, 89). Zur weiteren Verdeutlichung dieses Punktes habe ich die nachstehenden Gedanken Jacob von Uexkülls* wiedergegeben (aus dem von Thure von Uexküll herausgegebenen Buch KOMPOSITIONSLEHRE DER NATUR 1980):
p. 183: ...solange wir wie bisher ein jedes Tier mit unserer menschlichen Umwelt umkleiden, kommen wir aus den gröbsten Irrtümern gar nicht heraus. Nur von diesem falschen Standpunkt aus konnte man behaupten, die Lebewesen seien in einer dauernden Vervollkommnung begriffen, weil alle diejenigen Tiere, die weniger gut an die Natur angepaßt seien, ausstürben und nur die besser passenden überlebten.
Wenn man den Tieren ein unpassendes Menschengewand umwirft, gibt es freilich auch dazu unpassende Tiere. Erst wenn man die Tiere in ihr eigenes Umweltgewand kleidet, wird man gewahr, daß sie in dasselbe auf das vollkommenste eingepaßt sind.
- Worauf Beispiele folgen.
p. 281: Ein jedes Tier bildet den Mittelpunkt seiner Umwelt, der es als selbständiges Subjekt gegenübertritt...jede Umwelt eines Tieres bildet einen sowohl räumlich wie zeitlich, wie inhaltlich abgegrenzten Teil aus der Erscheinungswelt des Beobachters...Einen allgemeinen absoluten Raum und eine allgemeine absolute Zeit, die alle Lebewesen umschließen, gibt es nicht...
Von den Gesetzen, die das Leben schaffen und vernichten, können wir nur sagen, daß eine allumfassende Planmäßigkeit ihnen zugrunde liegt, die sich in der vollkommenen Einpassung eines jeden Lebewesens in seine Umwelt am deutlichsten ausspricht.
pp. 341/342: Wie die Zellen die elementaren Bausteine aller Lebewesen bilden, die nach bestimmten Bauplänen zusammengefaßt das Subjekt hervorbringen, das die Fähigkeit in sich trägt, eine Umwelt zu erbauen, so bilden die Umwelten die nächst höheren Bausteine, die, übersubjektiven Bauplänen gehorchend, das Gebäude der lebenden Natur errichten.
Diese grundlegende Erkenntnis der Lebewesen eröffnet uns den Ausblick in eine neue Mannigfaltigkeit, die bisher übersehen wurde. Eine Mannigfaltigkeit, die für den Mechanisten unerkennbar bleibt, weil er nur seine eigene Umwelt anerkennt und diese in der Vorstellung über alle Maßen ausgedehnt hat, um sie zum Tummelplatz für alle Lebewesen zu machen.
Doch ist diese eine Welt, mit nur einem Raum und einer Zeit, viel zu eng für das Gewimmel von Welten mit ihren abertausend Räumen und Zeiten, die unsere Augen blenden, sobald wir einmal den Blick über unsere eigene Umwelt erhoben haben, um das tausendfältige Weltgetriebe der Natur zu bewundern.
p. 370: Solange wir es nicht gelernt haben, die Umwelt eines Tieres aus der für unser Auge um das Tier ausgebreiteten Umgebung herauszuheben, machen wir uns eine völlig falsche Vorstellung von seinem Leben. Wenn wir das Tier in eine für unser Auge reiche und abwechslungsreiche Umgebung versetzen, die mit ihren tausend Reizen von allen Seiten auf das Tier einwirkt, werden wir nie fassen können, wie es möglich ist, daß die Tiere ihr Leben mit einer Sicherheit zu führen imstande sind, die allein mit dem unbeirrbaren Wachsen der Pflanzen zu vergleichen ist.
...Die Tiere nehmen von diesem Reichtum nichts wahr...Da zur Aufnahme von Reizen durch ein Tier entsprechende Sinnesorgane vorhanden sein müssen, verfährt die Natur ganz radikal, indem sie dem Tier keine Sinnesorgane außer den allernotwendigsten gewährt .
- Worauf zahlreiche Beispiele folgen, von denen wir nur das der Pilgermuschel zitieren möchten (p. 371):
Die Pilgermuschel, die hundert Augen aus ihrem Schalenrande hervorstreckt, aber ein äußerst primitives nervöses Zentralorgan besitzt, vermag keine Form, keine Farbe zu unterscheiden - einzig die langsame Fortbewegung ihres Spezialfeindes, des Seesternes, wird von ihr wahrgenommen. Jede andere Art von Bewegung bleibt unbemerkt. Außer dem Seestern ist kein anderer Bedeutungsträger in ihrer Umwelt vorhanden .
*Man kann ohne Schwierigkeiten Uexkülls Umweltlehre akzeptieren, ohne seinem philosophischen Idealismus zu folgen.
Entsprechend sind die Augen der Pilgermuschel (Pecten jacobaeus) auch für ihre Umweltbewältigung bereits so optimal gestaltet, dass sich eine Weiterentwicklung und "Vervollkommnung" erübrigt.
Diese Aussage gilt grundsätzlich für alle Lebensformen. Selbst Salvini-Plawen schreibt zu den Weichtieren 1979, p. 22:
Das Auftreten von Sinnesorganen richtet sich nach der Ausbildungsstufe, dem Lebensort und der Lebensweise der Tiere.**
Thure von Uexküll bemerkt zur Fortschrittsgläubigkeit des Darwinismus (1980, pp. 60/61):
Die Faszination, die der Entwicklungsgedanke ausgeübt hat und noch ausübt, ist nur so zu verstehen, wenn man die Analogie mit dem Fortschrittsgedanken sieht, der den Kern des Erlösungsglaubens der Industriekultur bildet. Fortschritt durch bessere technische Ausrüstung verspricht das Paradies auf Erden durch den Sieg über die Konkurrenz in der Ausbeutung der Natur. Der Darwinismus gibt diesen Glauben als eine "naturwissenschaftliche Wahrheit" aus.
- Und überträgt diesen Fortschrittsgedanken auf das gesamte Organismenreich.
**Salvini-Plawen und Mayr sprechen weiter (1977, p. 247) von "the little used and hence not subsequently modified "ancient" eyes of the Arcacea and Limidae"!
Kehren wir zu der Frage zurück, ob die Degenerationstheorie alle mit den verschiedenen Differenzierungsstufen des Auges verbundenen Probleme klärt, so können wir jetzt diese Frage mit einem klaren Nein beantworten. Wir müssten sonst sämtliche "niederen Lebensformen", mit ihren speziellen Innen- und Umwelten von den höher differenzierten und andersartigen Organismen ableiten. Die Theorie kann jedoch über eine Anzahl spezieller Fälle von Umweltverarmung und damit einhergehender Degeneration von Sinnesorganen innerhalb der primären Arten (die mehrere morphologische Gattungen bis hin zu verschiedenen Familien umfassen können - vgl. Lönnig 1988) Auskunft geben.
Wir möchten jetzt zu den einleitenden Behauptungen Salvini-Plawens und Mayrs, zitiert pp. 86/87, zurückkommen. Nach der oben schon wiedergegebenen Behauptung von einer "almost complete gradation", bei den Augen, schreiben die Autoren p. 209:
Indeed, the gradation from light-sensitive single cells through localized groups of such cells that serve as photoreceptive organs, to more complex organs (with focusing devices, etc.) is so continuous and complete that it became difficult to define what an eye is. Some authors have limited the term eye to the highly complex structures of the vertebrates, cephalopods, and arthropods, while others have freely applied the term eye to any photoreceptor organ beyond the one-cell stage. This fact alone tells the story of the evolution of eyes.
Auf die verschiedenen Diskontinuitäten in diesen Serien haben wir oben schon im Detail aufmerksam gemacht. Aber in diesem Absatz liegt wohl auch eine tiefe Wahrheit: Unsere begrifflichen und terminologischen Schwierigkeiten und Unklarheiten bilden die stärkste Evidenz für die Evolutionstheorie. Im übrigen liegt die Logik dieser Aussage auf derselben Ebene wie etwa die Behauptung, dass bei unterschiedlicher Terminologie einer Serie von immer tiefer werdenden Plastikschüsseln bis hin zum Eimer und weiter zum Abfalleimer diese Tatsache allein die Geschichte ihrer Evolution erzähle. Wir dürfen solche Beweise getrost in den Mülleimer der übrigen Evolutionsmythen werfen.
Weiter p. 209:
Thirdly, by applying population thinking to the study of variation of the eye, it has become quite evident that there is no such thing as "the eye" of a given species but that in the structure of the eye of a given species there is an enormous amount of variation among individuals, thus providing inexhaustible material for the operation of natural selection. There is, of course, only a single species, the human, in which this high variability has been studied intensively, but with our modern knowledge of the enormous genetic variability of all species there is no reason to doubt that a similar variability is universal in the animal kingdom.
Vgl. bitte zu diesem Missverständnis des Variationsthemas direkt die Ausführungen von Einhorn, zitiert pp. 42/43.
Wenn die Variabilität des menschlichen Auges ein "inexhaustible material for the operation of natural selection" bereitstellt, dann müsste es ein Leichtes sein, etwa ein Katzen-, Adler- oder Eulenauge beim Menschen zu entwickeln. Allen Untersuchungen und bekannten Tatsachen zufolge liegt eine solche Variabilität jedoch außerhalb der biologischen Realitäten. Verschwommene Begriffe bilden für weite Bereiche die Grundlagen der Synthetischen Evolutionstheorie. Und weil alle Begriffe beim Neodarwinismus zerfließen, kennt auch die Evolution keine Grenzen.
Die Realität: Neben einer großen neutralen Variabilität des gesunden menschlichen Auges, "der Unendlichkeit im kleinen", die jedoch immer in klaren Grenzen bleibt, sind weiter die schon p. 13 erwähnten genetisch bedingten pathologischen Veränderungen des Auges zu nennen. Die Letzteren werden jedoch meines Wissens von niemandem für eine weitere Vervollkommnung des menschlichen Auges eingesetzt. Nichtsdestoweniger ist den Autoren zuzustimmen, dass gründliche Untersuchungen zur genetischen Variation des Auges bei anderen Spezies die Probleme weiter erhellen könnten. Solche Studien dürften auch zusätzliche Beispiele für das Gesetz der Rekurrenten Variation* (Lönnig 1988) und damit die Möglichkeiten und Grenzen der Augenvariabilität bei den verschiedenen Spezies liefern.
*Die p. 43 zitierte "Unendlichkeit im kleinen" verteilt sich auf die begrenzte Anzahl der Mutantentypen.
Zum letzten Satz des vorigen Zitates kann man den Verfassern ebenfalls zustimmen (nur dass Strukturen, die bei anderen Lebensformen gar nicht auftreten, entsprechend auch nicht variieren können).
Weiter im Text von Salvini-Plawen und Mayr (pp. 209/210):
The two major misconceptions of Darwin's opponents were, first, to assume that the evolution of the eye had to be explained, defining "the eye" purely typologically, and, second, to ignore the actual process of natural selection by ascribing to Darwin a theory of the origin of eyes "by accident." Once these two misconceptions are removed, it becomes possible to interpret the rich modern information on the diversity of photosensitive organs in lower organisms strictly in terms of the Darwinian explanation, that is, as the result of an interaction between variation and selection.
Nur so kann man Darwin heute noch rechtfertigen: Man leugnet die biologische Realität des menschlichen Auges, indem man seine Variabilität ins grenzenlose schießen lässt, ohne zwischen neutraler und pathologischer Variabilität zu differenzieren.
Es kann sein, dass Darwins Opponenten die neutrale Variabilität in ihren Überlegungen nicht immer mit einbezogen haben. Sie waren jedoch mit ihrer "typologischen Betrachtungsweise" der Wahrheit immer noch näher als eine Ideologie, bei der sämtliche Begriffe verschwimmen und die die Wahrnehmung der Realität weitgehend blockiert. Außerdem ist die von Neodarwinisten regelmäßig vorgenommene Klassifizierung ihrer Kritiker als Typologen selbst eine Form der Typologie, die überdies völlig unfähig ist, die Variabilität in der kritischen Population wahrzunehmen.
"...second, to ignore the actual process of natural selection by ascribing to Darwin a theory of the origin of eyes "by accident"."
Wenn das wirklich ein realer Prozess wäre, der sich in der Vergangenheit zur Augenbildung vielfach abgespielt hat und sich auch heute noch überall abspielt, dann müsste er auch bei der zur Diskussion stehenden Frage reproduzierbar sein. Da die Selektion einer der beiden Schlüsselbegriffe zum Verständnis der gesamten Evolutionsfrage und damit auch der Entstehung des Auges sein soll, gebe ich hier über die schon p. 58 der vorliegenden Arbeit aufgeführten Punkte hinaus die pp. 466/467 aus meiner Artbegriffsarbeit (Lönnig 1988) wieder.
[Französische Biologen hatten folgenden Einwand zur Selektionstheorie erhoben:
Out of 120,000 fertilizes eggs of the green frog only two individuals survive. Are we to conclude that these two frogs out of 120.000 were selected by nature because they were the fittest ones; or rather... that natural selection is nothing but blind mortality which selects nothing at all?
(Vgl. vorliegende Schrift, p. 58)]
Die gleichen Fragen stellen sich für die (von Dobzhansky zitierten) 700 Milliarden Sporen von Lycoperdon, die 114 Millionen Eier mal der Zahl der Laichzeiten der Amerikanischen Auster, für die 28 Millionen Eier des Lachses usw. Salomo schrieb 1000 v.u.Z./1970, p. 688: "Wiederum sah ich unter der Sonne, dass nicht den Schnellen den Preis zufällt, und nicht den Helden der Sieg... sondern alle trifft Zeit und Zufall."
Es ist ja klar: Wenn von Millionen und Milliarden Individuen nur wenige (nämlich die zitierten 2 Nachkommen eines Elternpaars bei konstanter Population) überleben und zur Fortpflanzung kommen, dann ist es bei den unüberschaubaren Zufälligkeiten in der Natur gar nicht nachzuvollziehen, dass die beiden Tüchtigsten überleben sollen. Stark unterschiedliche Fähigkeiten zeichnen schon die Ontogenese aus. Beutetiere und Fressfeinde, Standortunterschiede und Witterungsbedingungen etc. gehören zu den ununterbrochen variierenden Parametern, so dass der Zufall bei dieser Frage ganz offensichtlich einen enormen Raum einnimmt. Es ist auch bei solchen Nachkommenzahlen viel wahrscheinlicher, dass statt der seltenen besten die 'mittelmäßigen' Rekombinanten überleben und sich fortpflanzen. Und die mittelmäßigen Individuen behaupten sich unter den jeweils gegebenen Umständen noch mit einer Vielzahl von neutralen bis schwach nachteiligen Genmutationen.
Wir wollen damit nicht sagen, dass es gar keine Selektion gibt. Selbstverständlich fallen physiologisch, anatomisch und ethologisch stark geschädigte Mutanten ihren Fressfeinden in der Regel schneller zum Opfer als normal funktionierende Rekombinanten. Auch werden solche Mutanten häufig Schwierigkeiten mit der Nahrungsbeschaffung haben. Nur auf Inselgebieten mit Ausfall oder Einschränkung solcher stabilisierender Selektion können stärkere Degenerationsprozesse schnell ablaufen. Anders sieht es jedoch mit den seltenen Rekombinanten aus, die in einer kontinuierlichen Evolution durch Mutationen mit 'geringer oder sogar unsichtbarer Wirkung auf den Phänotyp' die Anpassung und Höherentwicklung besorgen sollen. Das müssten ja ausgesprochen seltene Mutanten und Rekombinanten sein, die den Rahmen der rekurrenten Variation überschreiten. Bei den oben zitierten Zahlen hätten jedoch solche Individuen kaum eine Chance, zur Fortpflanzung zu gelangen, - abgesehen davon, dass es Schwierigkeiten mit der Beschaffung eines entsprechend weiterentwickelten Partners geben dürfte. Dazu kommt noch, dass die aufgrund wechselnder Umweltverhältnisse von Individuum zu Individuum auftretenden nichtgenetischen Phänotypunterschiede die nahezu unsichtbaren Auswirkungen der Mikromutation bei weitem übertreffen können. Vielleicht könnte in ganz seltenen Fällen hier noch der von neodarwinistischer Seite für die Selektion so stark verneinte Zufall weiterhelfen. Aber dafür dürfte die Wahrscheinlichkeit so gering sein, dass schon ein starker Optimismus notwendig wäre, um damit generell den Ursprung der Lebensformen zu erklären.
Ein weiterer Punkt: Da die Unterschiede in der Zahl der Nachkommenschaft z.B. zwischen einem Hering und einer Elefantenkuh nicht zu übersehen sind, können auch die Gametenproduktion und Nachkommenzahl nicht einfach als gegeben angesehen werden. Worauf gehen die Unterschiede zurück? Nach neodarwinistischer Auffassung sind sie letztlich auch wieder auf die alle Strukturen und Fähigkeiten erzeugenden Zufallsmutationen zurückzuführen. Wenn es also die aufgrund der Nachkommen-Überproduktion postulierte übergeordnete Selektion gäbe, dann wäre sie selbst auch wieder ein Produkt der Mikromutationen!
Setzen wir aber einmal die alles überwaltende Selektion im Sinne der Synthetischen Evolutionstheorie voraus: Worauf beruht das "survival of the fittest"? Das Überleben ist ganz klar abhängig von der Funktionsfähigkeit der in den Organismen zusammenwirkenden Strukturen, nach deren Ursprung wir oben fragten. Ein Hase läuft schneller, ein Löwe springt weiter, ein Zebra wittert besser, ein Adler sieht schärfer, ein Schimpanse reagiert genauer etc. als seine Artgenossen, weil nach neodarwinistischer Lehre die Zufallsprozesse der Mutation und Rekombination sie in einer kontinuierlichen Evolution mit allen bisherigen Strukturen und auch den neuerlichen Verbesserungen ausgerüstet haben. "Chance" events bestimmen damit alles: Form und Funktion der Strukturen in allen Bereichen, diese die natürliche Auslese im Kampf ums Dasein und damit die gesamte biologische Evolution. Ohne Form und Funktion bereits vorhandener Strukturen gibt es keine Selektion. Die Selektion ist damit auch nicht "self-steering" und ähnliches mehr, sondern die Folge der durch "chance" events entstandenen Formen und Fähigkeiten, samt Überproduktion der Nachkommenschaft. (Vgl. auch Endler 1986.) Die gesamte Organismenwelt ist damit auf Zufallsereignisse zurückzuführen. Diese Theorie ist ihrem tiefsten Wesen nach eine Zufallslehre und die Kritiker treffen den Kern der Sache, wenn sie unter anderen Fragen diesen Punkt ins Visier nehmen.
Kommen wir im Anschluss an diese Ausführungen zurück zu Salvini-Plawens und Mayrs Beitrag zur Entstehung des Auges. Sie hatten ihren Kritikern vorgeworfen, 'den tatsächlichen Prozeß der natürlichen Selektion' bei der Bildung des Auges zu ignorieren etc. - Worin besteht nun nach diesen Autoren der Selektionsprozess bei der Entstehung von Lichtsinnesorganen und Augen? Neben ein paar generalisierenden Behauptungen auf p. 251 ihrer Arbeit, schreiben die Autoren zur konkreten Frage p. 249:
Precisely what the selection pressures are that lead to the various types of differentiated photoreceptors is insufficiently understood. There seem to be three basic functions of eyes, simply to detect changes in the amount of incident light (light/shadow detection), to determine the direction of the incident light (orientation), or to analyze the environment through image formation. Since we have very little understanding of visual perception in almost all invertebrate groups we are unable to attribute the capacity for image formation to a certain structure of eyes except for inference from the arrangement of the elements (see Pecten; Land, 1968, p. 81; Weale, 1974).
Wir können immer nur den (in aller Regel zufriedenstellenden) Funktionswert der Augen der verschiedenen Arten für ihre Umwelt und neurale Organisation feststellen. Gehen wir jedoch ins Detail bei der Frage nach den über Tod und Leben entscheidenden Selektionswerten der postulierten Mikroevolutionsprozesse, dann geraten wir regelmäßig in das Dilemma von funktionsuntauglichen Zwischenformen, die dabei aber gleichzeitig einen so entscheidenen Selektionsvorteil gehabt haben sollen, dass sie alle ihre Verwandten in den Schatten der Vergangenheit stellen konnten (vgl. p. 58, Absatz X). Dabei sehen wir vom Synorganisationsproblem und allen damit verbundenen mathematischen Unwahrscheinlichkeiten zufälliger Mutationsprozesse noch einmal ganz ab.
Der Selektionsdruck kann jedoch in solchen Phasen wesentlich besser wie folgt verstanden werden: Wenn es ihn in der von Darwin und den Neodarwinisten geforderten Form gibt, dann hat er - als Prozess, der aufmerksam jede geringe Veränderung beobachtet und mit absoluter Sicherheit nur die Besseren erhält - all diesen funktionsuntauglichen Zwischenformen auch millionenfach ein Ende bereitet und damit eine etwaige "Vervollkommnung" und jede weitere Evolution vollständig blockiert.
Im Zitat von p. 104 lasen wir:
Once these two misconceptions are removed, it becomes possible to interpret the rich modern information on the diversity of photosensitive organs in lower organisms strictly in terms of the Darwinian explanation, that is, as the result of an interaction between variation and selection.
Nach unseren bisherigen Ausführungen liegen - klar erwiesen - die Missverständnisse auf darwinistischer Seite. Folglich ist es nicht einmal mehr möglich, die Entstehung der verschiedenen Augentypen widerspruchsfrei mit darwinistischen Erklärungsprinzipien zu interpretieren.
Um einen postulierten Prozess in den Rang einer naturwissenschaftlichen Tatsache zu erheben - wie das die Autoren im Laufe ihrer Arbeit mit der Evolutionstheorie immer wieder tun - reicht es prinzipiell nicht aus, ein bestimmtes Faktenmaterial im Sinne einer bestimmten Auffassung, hier des evolutionistischen Weltbildes, interpretieren zu können.
Um noch einen besseren Maßstab zur Bewertung der folgenden Behauptungen aus der abschließenden Diskussion und Zusammenfassung der Autoren in der Hand zu haben, müssen wir uns noch vergegenwärtigen, dass sie in ihrer Abhandlung über 90% aller komplexen Strukturen und Prozesse des menschlichen Auges und Gehirns (oder der Wirbeltiere schlechthin) überhaupt nicht erwähnen, und darüber hinaus auch das Synorganisationsproblem völlig ausklammern.
In der Schlussdiskussion p. 253 und der Zusammenfassung p. 254 lesen wir unter anderem:
The original purpose of our investigation had been to test the validity of Darwin's assertion that the evolution of eyes is no stumbling block for his theory of natural selection. As we have seen, Darwin passed this test with flying colors.
The structure that was forever cited by Darwin's opponents as one "which could not possibly have been formed by numerous successive, slight modifications" was the eye. In much of the anti-Darwinian literature it became the test object of natural selection. Although Darwin himself had already advanced cogent arguments against the validity of these objections, there has been no comprehensive treatment of this problem since Hesse (1896-1908) and Plate (1924).
The results of our analysis completely substantiate Darwin's claims, but also reveal numerous still unsolved problems.
Mit der Umformulierung zwecks Richtigstellung der anfangs (pp. 87/88) und weiter unten (p. 105 ff.) zitierten Ausführungen Salvini-Plawens und Mayrs möchten wir die Hauptergebnisse unserer Untersuchungen wie folgt zusammenfassen:
The modern evidence showing that Darwin was wrong is overwhelming. First of all, it is now quite obvious that the evolution of an organ of vision is vastly more improbable than was thought in Darwin's day. Electron microscopy and other modern methods of investigation have shown that the structures of photoreceptors as well as their biochemical functions are much more complex than could have been surmised a hundred years and more ago. Moreover, photoreceptors of various degrees of differentiation have appeared independently of each other in at least 40, if not 65 or even more separate animal groups. As the hypothesis regarding their origin by micromutations and selection is already so highly improbable that it is irreproducible, a 65-times independent origin by the same factors is an evolutionary myth far beyond the boundaries of modern science.
Secondly, there exists in living animals no complete gradation from exceedingly simple light-sensitive receptors to highly complex eyes. The fact that different specialists use different terms for various types of eyes has, of course, nothing to do with the mode of their origins.
Thirdly, considering the eye's "sheer number of interlocking parts, all well-adapted to seeing and well-adapted to each other" in man (neo-Darwinian Dawkins) as well as applying population thinking to the study of the variation of the eye and its functional relations to the organism as a whole and its environment, it has become evident that obviously there exists such an integrated system as "the eye" for a given species. Although there is an enormous amount of neutral variation in the structure of the eyes among individuals, it never passes the genetical, developmental and functional boundaries of the synorganized system except in pathological cases, thus never providing the inexhaustible material so desperately needed for the operation of natural selection and evolution to generate entirely new and more highly differentiated types of eyes.
There is, as yet, only a single species, the human, in which the neutral and pathological variability has been studied intensively. But with our modern knowledge of the neutral variability of all species, which is, however never transcending the boundaries of their basic types and/or never generating new synorganized anatomical and physiological systems, there is now doubt that a similar variability is universal in the animal kingdom: because of the improbability of a large number of coordinated series of information-generating, independent random micromutations, no really complex states of eyes can have evolved from simple ones by mutations and selection. (See also the Law of Recurrent Variation, Lönnig 1988.)
The two major correct conceptions of Darwin's opponents were, first to assume that there are such synorganized systems of the eye with genetical, developmental and functional boundaries of variation and second to question the hypothetical process of natural selection, ascribing to Darwin a theory of the origin of eyes mainly by accident, especially in the non-functional intermediate links. Once the two neo-Darwinian misconceptions of limitless variation and positive selection values for all the necessary millions of microevolutionary intermediate links are removed, it becomes reasonable and necessary to interprete the rich modern information on the diversity of photosensitive organs in lower and higher organisms in terms of creation, that is, as the result of thoughtful, ingenious and deliberate design.
The construction of living organisms is not the result of selected variations, but variation is an inherent part of the construction.
The original purpose of my investigation had been to impartially test the validity of Darwin's assertion that a random evolution of eyes is no stumbling block for the theory of natural selection. As we have seen, Darwin failed completely. As the evolutionist of today must omit most of the complex eye structures and -synorganizations of vertebrates in order to persuade their followers in the absence of scientific facts, and as excessive complexities undreamt of in Darwin's days have been discovered by modern methods of electron microscopy and biochemistry, - the eye as a structure that has always been cited by Darwin's opponents as one "which could not possibly have been formed by numerous successive slight modifications" has become an even better example against the Darwinian and neo-Darwinian errors and misconceptions than ever before. Thus, in much of the anti-Darwinian literature it has convincingly become one of the core arguments against the hypothesis of the origin and evolution of all life-forms by micromutations and natural selection. Darwin himself had already doubted his own arguments as he was unable to advance any cogent points against the validity of the objections of his opponents.
The result of the present analysis completely disprove Darwin's claims, but also reveal numerous still unsolved problems.